
红树林湿地碳储存和主要有机碳过程研究进展
|
靳瑜(1993—),山西省宁武人,博士研究生,从事湿地生态研究。E-mail: Yu_Jin93@163.com |
收稿日期: 2024-11-25
修回日期: 2025-05-15
网络出版日期: 2026-03-12
版权
Advances in carbon storage and main organic carbon processes in mangrove wetlands
Received date: 2024-11-25
Revised date: 2025-05-15
Online published: 2026-03-12
Copyright
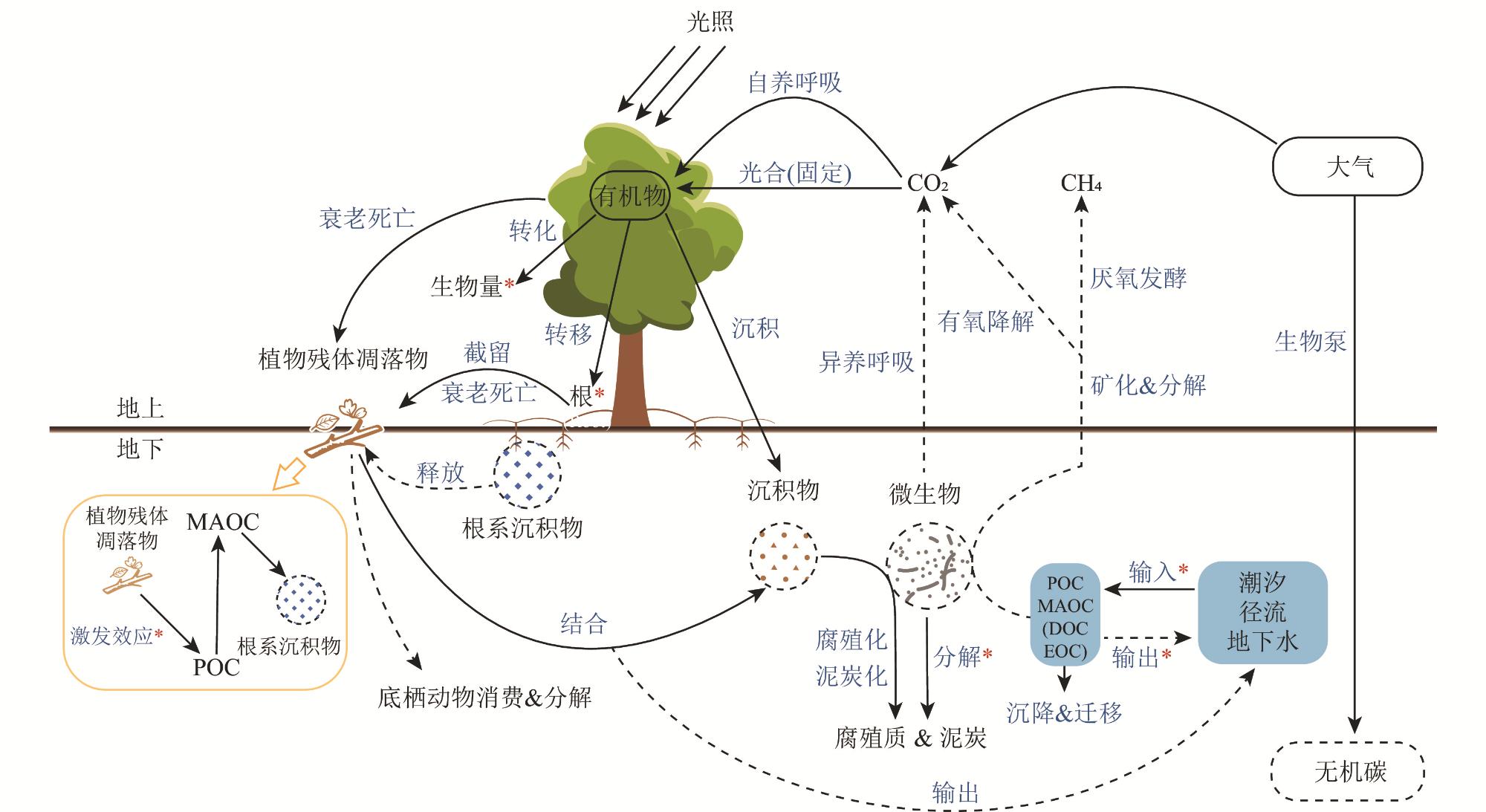
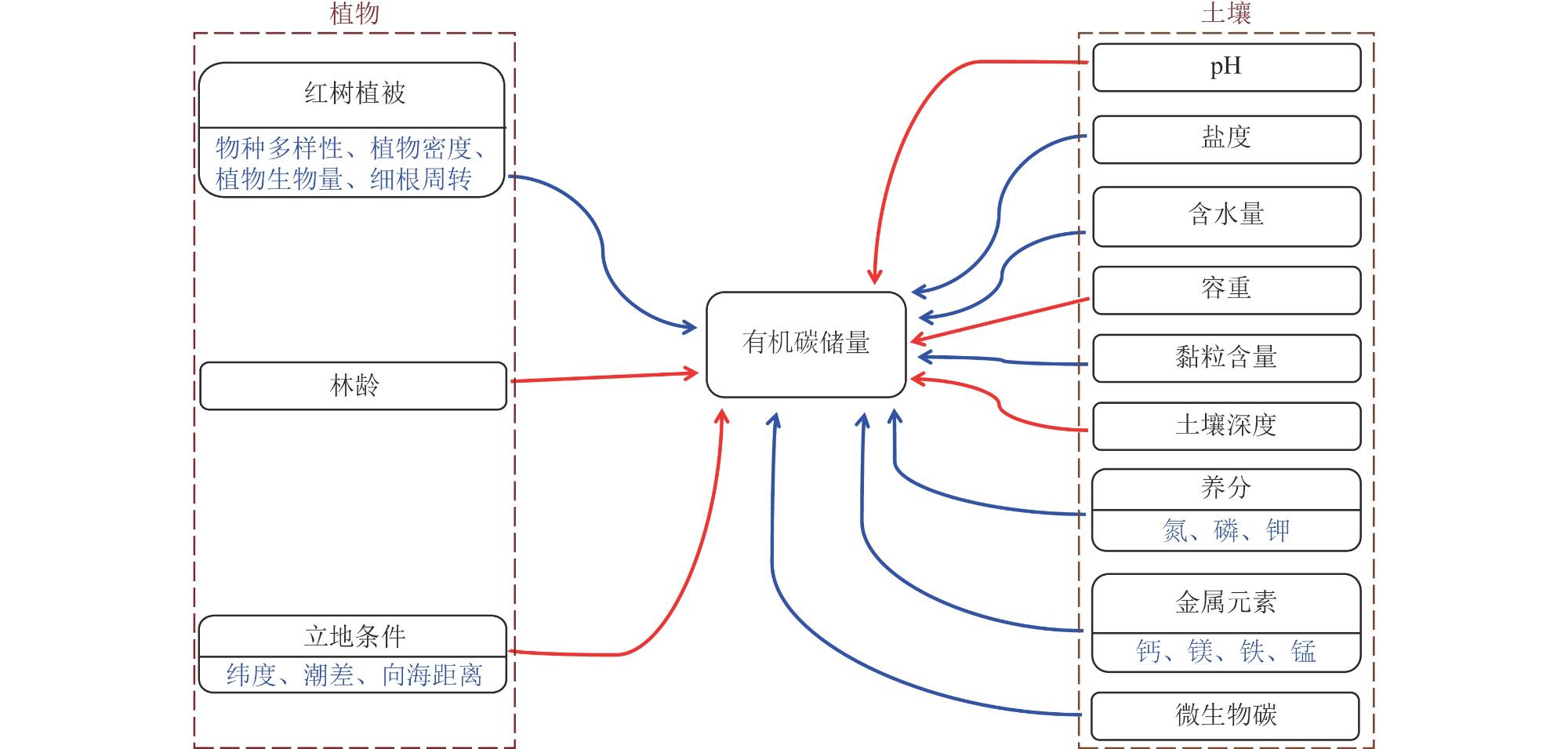
红树林湿地作为海洋与陆地之间的天然屏障,对大气中CO2、海洋环境中有机碳的捕获及碳固持具有重要作用。红树林湿地中的碳最终以植被生物量、植物凋落物和土壤有机碳的形式固存于生态系统内,有机碳的输出主要以气体形态(CO2和CH4)、底栖动物取食以及水体输出的方式损耗。本文通过归纳总结2018—2023年相关文献的研究结果,对红树林湿地碳储存(碳储量、碳密度和碳固存速率)及其影响因素(植物、土壤理化性质等)、红树林湿地碳循环主要有机碳过程(碳输入和碳输出)的研究进展进行了综述,提出了有效恢复并提升红树林碳汇能力、平衡保护与修复的关系是目前红树林湿地修复过程中亟待解决的问题。建议未来加强红树林湿地碳过程的影响因素、碳库转化机制、植被修复效益、碳汇的恢复过程、激发效应及消费者(底栖动物)在碳过程中的作用等方面的研究,从而为红树林湿地的保护与修复、缓解温室效应以及达成“双碳”目标提供科学参考。
靳瑜 , 郭嘉 , 赵微 , 吴婷 , 王荣 , 林广旋 , 崔圆 , 罗芳丽 , 朱耀军 . 红树林湿地碳储存和主要有机碳过程研究进展[J]. 湿地科学, 2026 , 24(1) : 224 -234 . DOI: 10.13248/j.cnki.wetlandsci.20240295
As a critical natural ecotone and barrier between marine and terrestrial ecosystems, mangrove wetlands play an indispensable role in the capture and sequestration of atmosphere carbon dioxide (CO2), as well as organic carbon in the marine environment. Carbon within mangrove wetland ecosystems is ultimately fixed through a series of biological processes and biogeochemical reactions and sequestered in three primary and stable forms: vegetation biomass (i.e., plant aboveground and belowground biomass), plant litters (such as leaves, branches and root, etc.), and soil organic carbon (SOC). In contrast, the export and emission of organic carbon from mangrove wetlands occur mainly through three dominant pathways: the fluxes of greenhouse gas (mainly CO2 released via soil respiration and litter decomposition, and methane (CH4) produced under anaerobic soil conditions), the feeding and activities of benthic fauna (such as crabs and snails, etc.), and the hydrological export of dissolved organic carbon (DOC) and particulate organic carbon (POC) into adjacent coastal waters via tidal flow and runoff. By synthesizing the research findings reported in relevant domestic and international publications over the past six years (2018—2023), we reviewed the advances in three key processes of mangrove wetland carbon cycling, including carbon sequestration (i.e., carbon storage, carbon density and carbon sequestration rate) and its influencing factors (e.g., plant, soil physical and chemical properties), as well as the main organic carbon processes (i.e., carbon input and carbon export) in the carbon cycle of mangrove wetlands. Based on the review, we propose two urgent scientific and practical issues that need to be addressed in the current ecological restoration of mangrove wetlands. The first issue is the restoration and enhancement of the carbon sink capacity of degraded mangrove wetlands. The second issue is the need to balance the relationship between conservation (which have high and stable carbon sink capacities and should be protected from disturbance) and the ecological restoration of degraded mangrove areas. To address these gaps, future research should: in-depth exploration of the interactive effects of biotic and abiotic factors on mangrove carbon processes; systematic investigation of the transformation mechanisms of carbon pools; quantitative evaluation of the carbon sink benefits of vegetation restoration; detailed evaluation of restoration processes of carbon sinks; research on the priming effects and roles of consumers (e.g., benthic fauna) in carbon dynamics. Our review could provide a scientific support for the protection and restoration of mangrove wetlands, mitigation of greenhouse effect, and achieving the goals of carbon emissions peaking and carbon neutrality.
| 1 |
| 2 |
| 3 |
| 4 |
| 5 |
| 6 |
| 7 |
| 8 |
| 9 |
| 10 |
| 11 |
| 12 |
| 13 |
| 14 |
| 15 |
| 16 |
| 17 |
| 18 |
| 19 |
| 20 |
| 21 |
| 22 |
| 23 |
| 24 |
| 25 |
| 26 |
| 27 |
| 28 |
| 29 |
| 30 |
| 31 |
| 32 |
| 33 |
| 34 |
| 35 |
| 36 |
| 37 |
| 38 |
| 39 |
| 40 |
| 41 |
| 42 |
| 43 |
| 44 |
| 45 |
| 46 |
| 47 |
| 48 |
| 49 |
| 50 |
| 51 |
| 52 |
| 53 |
| 54 |
| 55 |
| 56 |
| 57 |
| 58 |
| 59 |
| 60 |
| 61 |
| 62 |
| 63 |
| 64 |
| 65 |
| 66 |
| 67 |
| 68 |
| 69 |
| 70 |
| 71 |
| 72 |
| 73 |
| 74 |
| 75 |
| 76 |
| 77 |
| 78 |
| 79 |
| 80 |
| 81 |
| 82 |
| 83 |
| 84 |
| 85 |
| 86 |
| 87 |
| 88 |
| 89 |
| 90 |
| 91 |
| 92 |
| 93 |
| 94 |
| 95 |
| 96 |
| 97 |
| 98 |
| 99 |
| 100 |
| 101 |
| 102 |
| 103 |
| 104 |
| 105 |
| 106 |
| 107 |
| 108 |
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}