
氮添加对大叶章生长和无性繁殖的影响
|
王梦雨(1998—),女,河南省柘城人,硕士研究生,从事湿地生态学研究。E-mail: 2452290847@qq.com |
收稿日期: 2024-10-18
修回日期: 2025-02-16
网络出版日期: 2026-03-12
版权
Effects of nitrogen addition on the growth and asexual reproduction characteristics of Deyeuxia purpurea
Received date: 2024-10-18
Revised date: 2025-02-16
Online published: 2026-03-12
Copyright
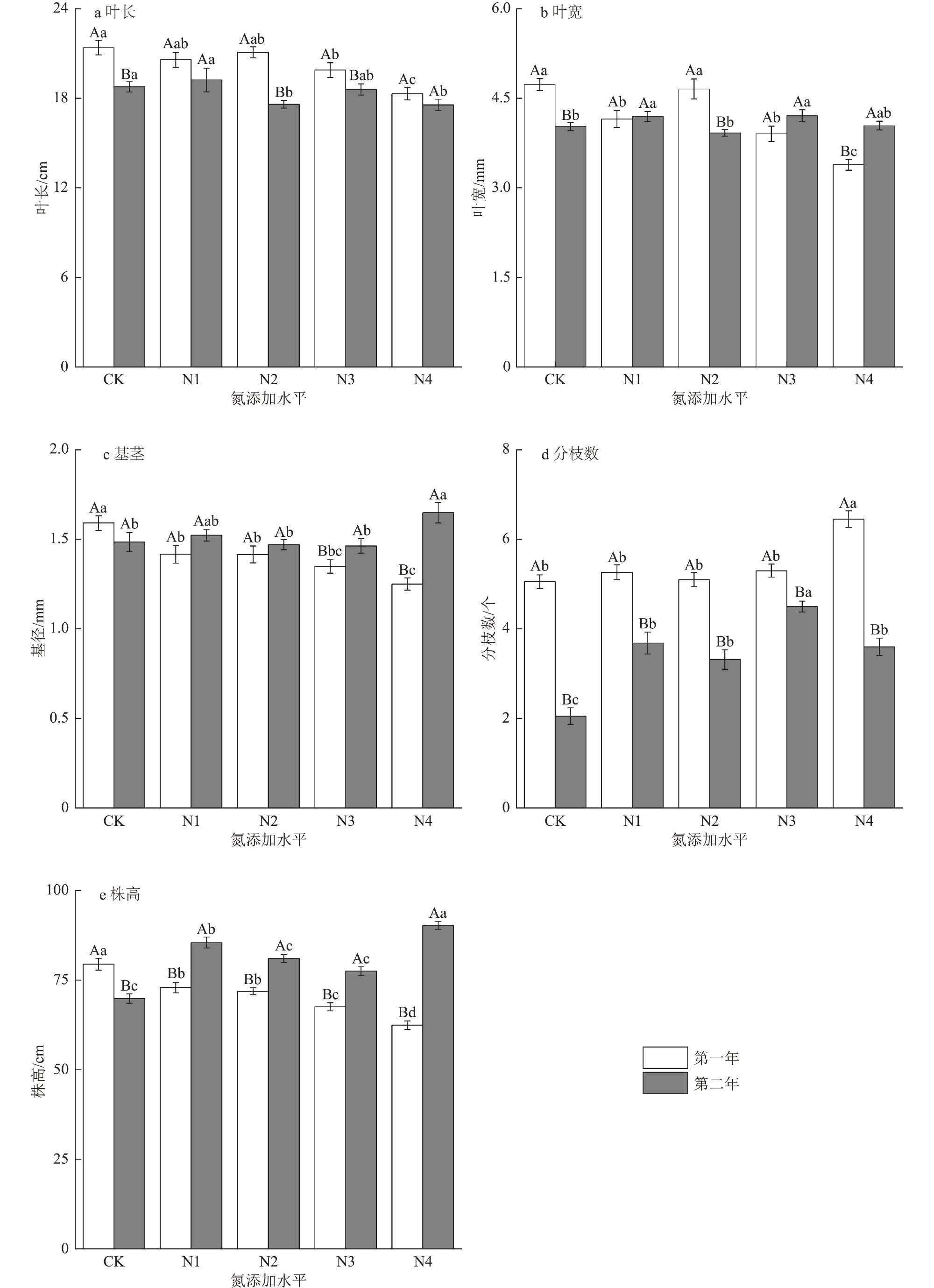
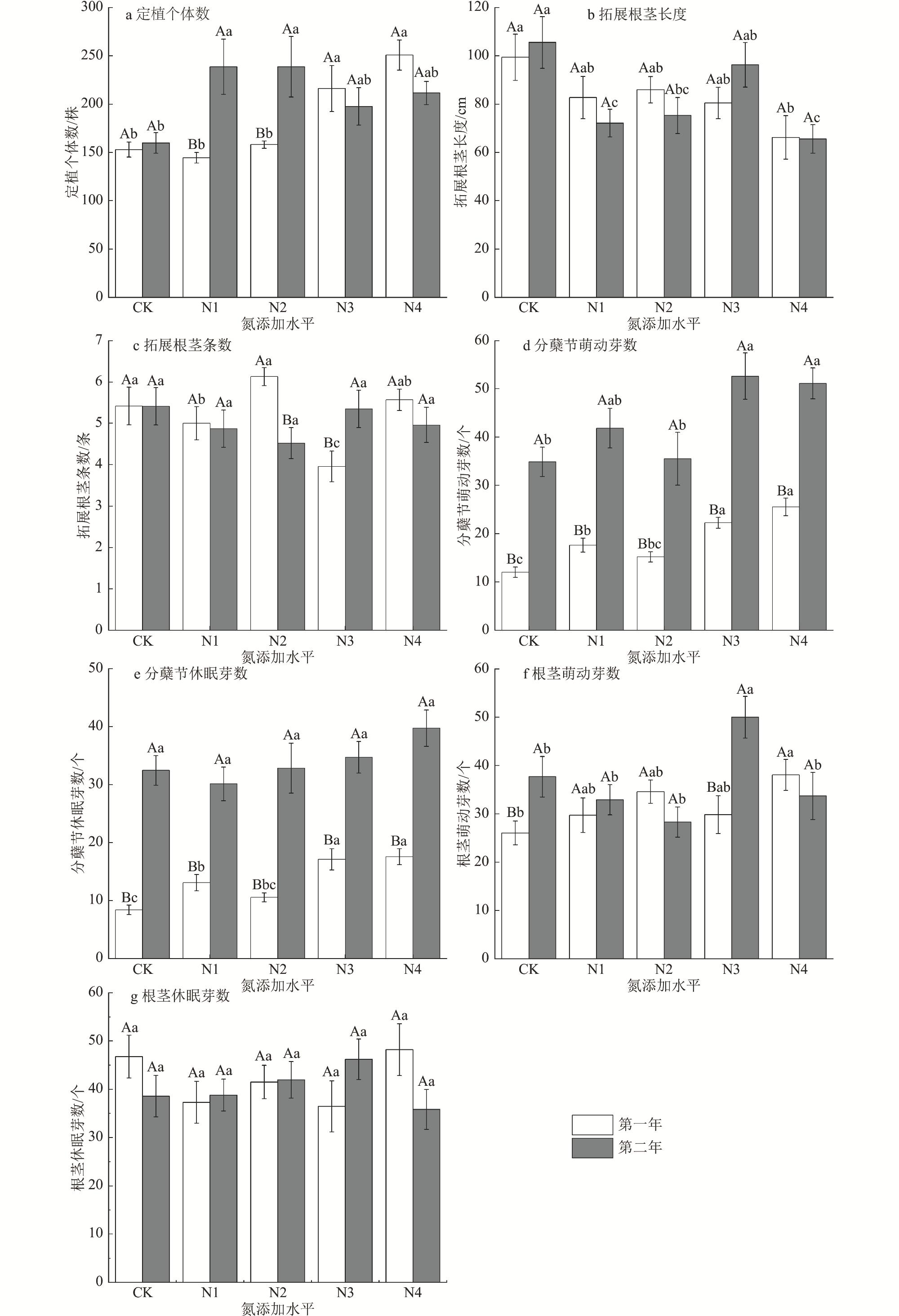
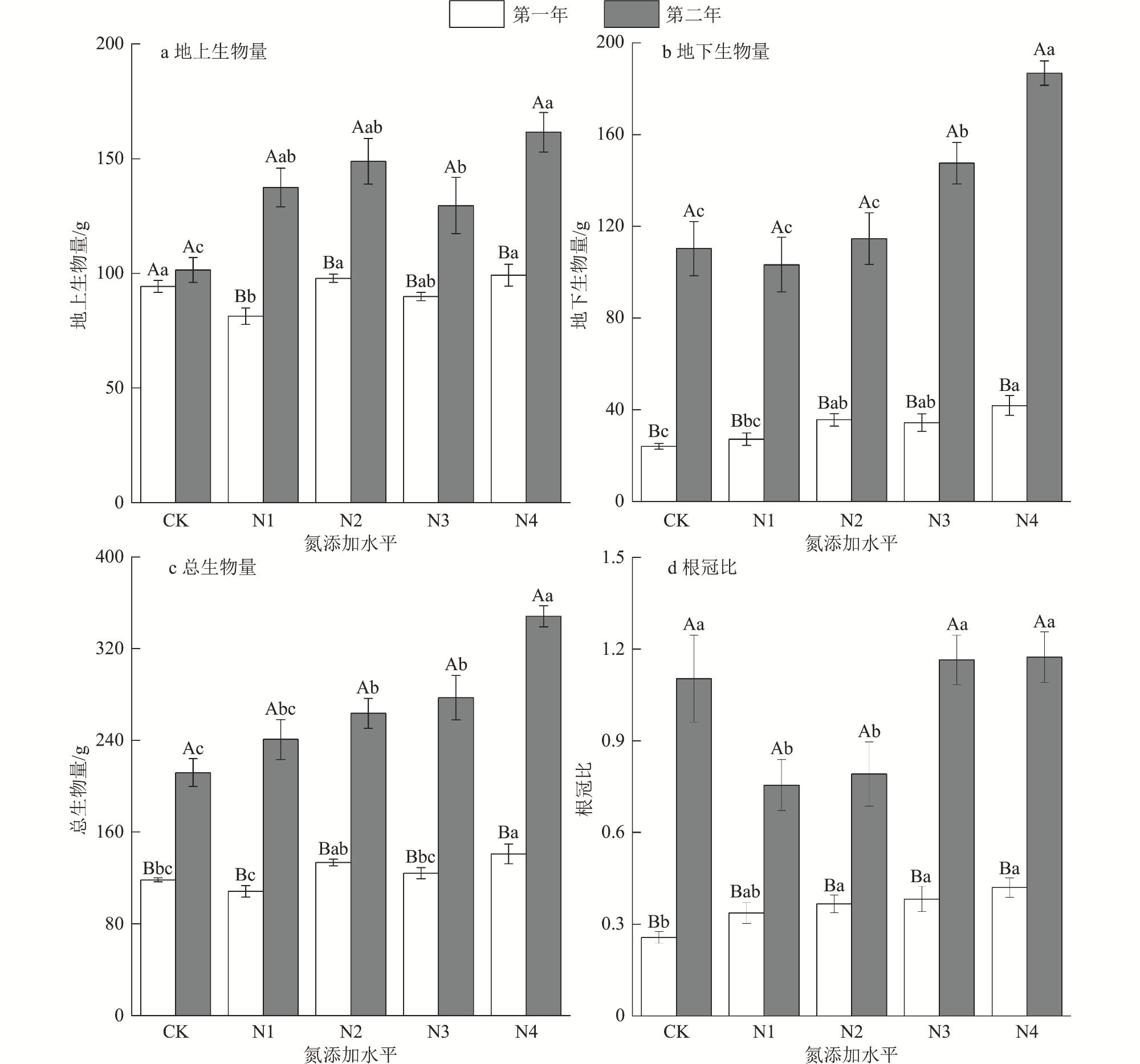
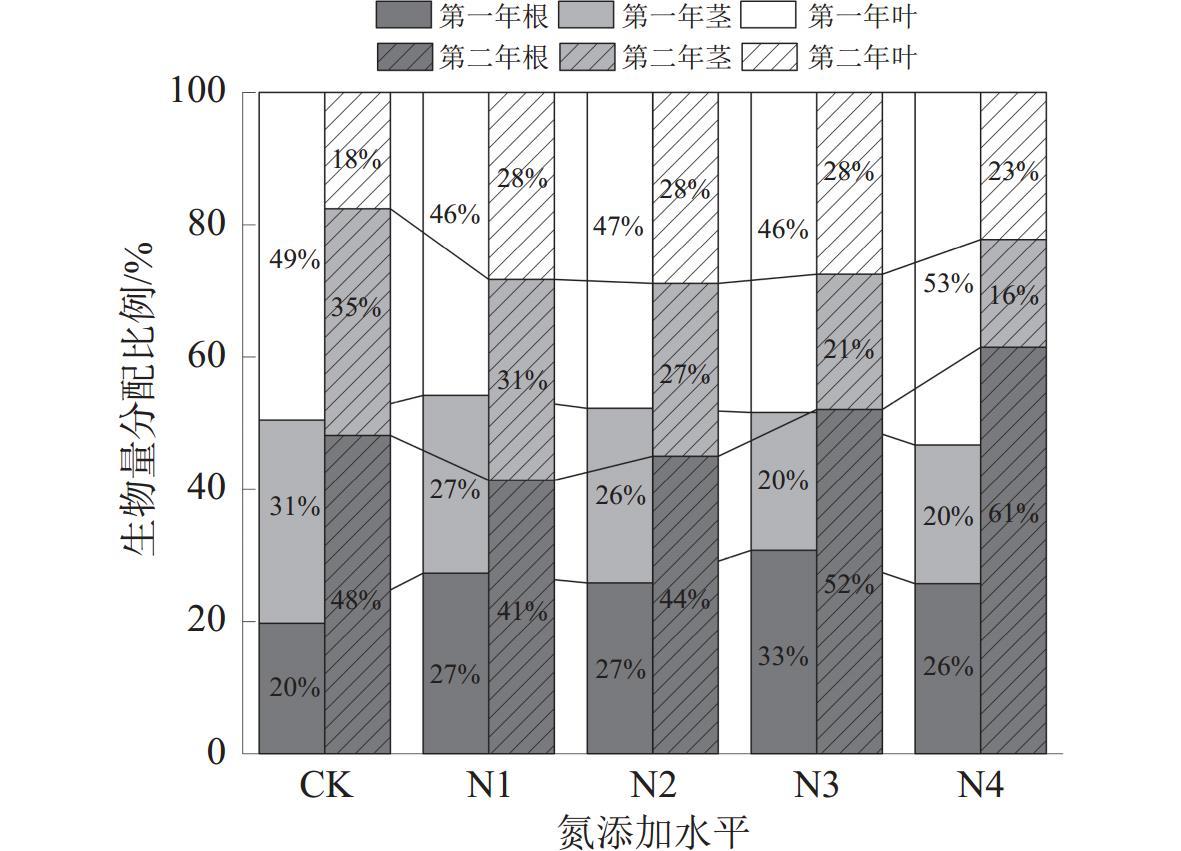
为了揭示湿地植被对氮沉降的响应机制,以三江平原湿地优势植物大叶章(Deyeuxia purpurea)为研究对象,通过控制实验研究了氮添加对其功能性状、无性繁殖和生物量分配的影响。试验设置CK[0 g/(m2·a)]、N1[2 g/(m2·a)]、N2[4 g/(m2·a)]、N3[6 g/(m2·a)]和N4[8 g/(m2·a)]5种氮添加水平,试验周期为2 a。研究结果表明,氮添加时间和水平及二者交互作用对大叶章株高、基茎、分株数有极显著影响(p<0.01),氮添加增加了大叶章分枝数量,降低了叶长;氮添加第一年显著降低了株高和基茎(p<0.01),第二年则呈相反趋势。氮添加时间和水平对大叶章分蘖节萌动芽和休眠芽数有显著影响(p<0.01),其交互作用对定植个体数、拓展根茎条数和根茎萌动芽数有极显著影响(p<0.01)。氮添加增加了定植个体数、分蘖节萌动芽数和休眠芽数,降低了拓展根茎长度和根茎条数。随着氮添加量的增加,大叶章总生物量、地上生物量和地下生物量逐渐增加,且氮添加第二年显著高于氮添加第一年(p<0.01),根和叶生物量分配比例逐渐增加而茎的分配比例减少。氮添加增加了大叶章分枝数、分蘖节芽数、根茎萌动芽数和定植个体数,降低了叶长,减少了拓展根茎条数和长度,促进了生物量积累,氮添加后大叶章会通过“密集型”生长策略扩大种群数量。
王梦雨 , 李旭 , 孙杰 , 曹宏杰 , 徐明怡 , 付晓玲 , 刘赢男 . 氮添加对大叶章生长和无性繁殖的影响[J]. 湿地科学, 2026 , 24(1) : 177 -188 . DOI: 10.13248/j.cnki.wetlandsci.20240280
Under the background of global change, to explore the mechanism of wetland vegetation response to nitrogen deposition, we selected Deyeuxia purpurea which is one of the dominant plants of Sanjiang Plain Wetland as the research object, through controlled experiments to study the effects of nitrogen addition level and nitrogen addition time on its functional traits, asexual reproduction and biomass allocation. In this controlled experiment, five nitrogen addition levels were set, CK[0 g/(m2·a)], N1[2 g/(m2·a)], N2 [4 g/(m2·a)], N3[6 g/(m2·a)]and N4[8 g/(m2·a)], and the experimental period was 2 years. The research results indicated that nitrogen addition time, nitrogen addition level and their interaction had extremely significant impacts on plant height, base diameter, and number of branches of D. purpurea (p<0.01). Nitrogen addition increased the number of branches, decreased the leaf length, and significantly reduced the plant height and base diameter of D. purpurea in the first year (p<0.01), while showed opposite trends in the second year. Nitrogen addition time and levels had significant effects on the number of dormant buds and sprouting buds at the tillering nodes of D. purpurea (p<0.01), and their interaction had extremely significant effects on the number of established individuals, the number of elongating rhizome branches, and the number of sprouting buds of rhizomes (p<0.01). Nitrogen addition increased the number of established individuals, the number of sprouting buds at tillering nodes, and the number of dormant buds, but decreased the length and number of elongating rhizome branches. Total biomass, above-ground biomass, and below-ground biomass of D. purpurea gradually increased with the increase of nitrogen addition concentration, and the results of the second year were significantly higher than those of the first year (p<0.01). The allocation proportion of root and leaf biomass gradually increased, while the allocation proportion of stem biomass decreased. Our study demonstrated that the addition of nitrogen increased the number of branches, tillering buds, sprouting buds of rhizomes, and the number of established individuals of D. purpurea, decreased the leaf length, the number and length of elongating rhizome branches, and promoted the biomass accumulation. At nitrogen addition levels of 6 g/(m2·a) and 8 g/(m2·a), and with an additional period of two years, it effectively regulated the growth and asexual reproduction of D. purpurea. The results showed that D. purpurea through an ‘intensive’ growth strategy to expand its population number under the the nitrogen addition background.
1 Two-factor analysis of variance results of effects of nitrogen addition time and levels on functional traits of Deyeuxia purpurea氮添加时间和水平对大叶章功能性状影响的双因素方差分析结果 |
| 处理 | df | 株高 | 叶长 | 叶宽 | 分枝数 | 基径 | |||||||||
| F | p | F | p | F | p | F | p | F | p | ||||||
| 时间 | 1 | 151.218 | <0.001 | 42.678 | <0.001 | 1.623 | 0.204 | 304.999 | <0.001 | 16.758 | <0.001 | ||||
| 氮添加 | 4 | 17.396 | <0.001 | 6.771 | <0.001 | 11.254 | <0.001 | 21.163 | <0.001 | 4.380 | 0.002 | ||||
| 时间×氮添加 | 4 | 43.320 | <0.001 | 2.960 | 0.021 | 16.164 | <0.001 | 12.844 | <0.001 | 4.814 | 0.001 | ||||
2 Two-factor analysis of variance results of effects of nitrogen addition time and levels on asexual reproduction characteristics of Deyeuxia purpurea氮添加时间和水平对大叶章无性繁殖特征影响的双因素方差分析结果 |
处理 | df | 定植个体 数 | 拓展根茎 条数 | 拓展根茎 长度 | 分蘖节 萌动芽数 | 分蘖节 休眠芽数 | 根茎 萌动芽数 | 根茎 休眠芽数 | |||||||||||||
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||||||||
| 时间 | 1 | 4.405 | 0.038 | 0.595 | 0.441 | 0.001 | 0.998 | 154.678 | <0.001 | 174.984 | <0.001 | 4.585 | 0.033 | 0.424 | 0.516 | ||||||
| 氮添加 | 4 | 4.594 | 0.005 | 1.274 | 0.281 | 4.036 | 0.003 | 9.425 | <0.001 | 3.957 | 0.004 | 2.241 | 0.065 | 0.347 | 0.846 | ||||||
| 时间×氮添加 | 4 | 5.310 | 0.002 | 3.794 | 0.005 | 2.701 | 0.031 | 0.708 | 0.587 | 0.798 | 0.527 | 4.787 | 0.001 | 2.029 | 0.091 | ||||||
3 Two-factor analysis of variance results of nitrogen addition time and levels on biomass of Deyeuxia purpurea氮添加时间和水平对大叶章生物量的双因素方差分析结果 |
| 处理 | df | 总生物量 | 地上生物量 | 地下生物量 | 根冠比 | |||||||
| F | p | F | p | F | p | F | p | |||||
| 时间 | 1 | 421.376 | <0.001 | 99.079 | <0.001 | 437.99 | <0.001 | 183.167 | 0.005 | |||
| 氮添加 | 4 | 15.662 | <0.001 | 6.916 | <0.001 | 14.68 | <0.001 | 4.445 | <0.001 | |||
| 时间×氮添加 | 4 | 7.174 | <0.001 | 5.05 | 0.002 | 7.460 | <0.001 | 3.756 | 0.011 | |||
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}