
基于生物量概念的互花米草时空分布元胞自动机模型
|
蔡晗(2000—),男,河南省许昌人,硕士研究生,从事植被水流相互作用模拟研究。E-mail: 22234165@zju.edu.cn |
收稿日期: 2024-08-16
修回日期: 2025-01-22
网络出版日期: 2026-03-12
版权
Cellular automaton model of Spartina alterniflora spatial-temporal distribution based on biomass concept
Received date: 2024-08-16
Revised date: 2025-01-22
Online published: 2026-03-12
Copyright
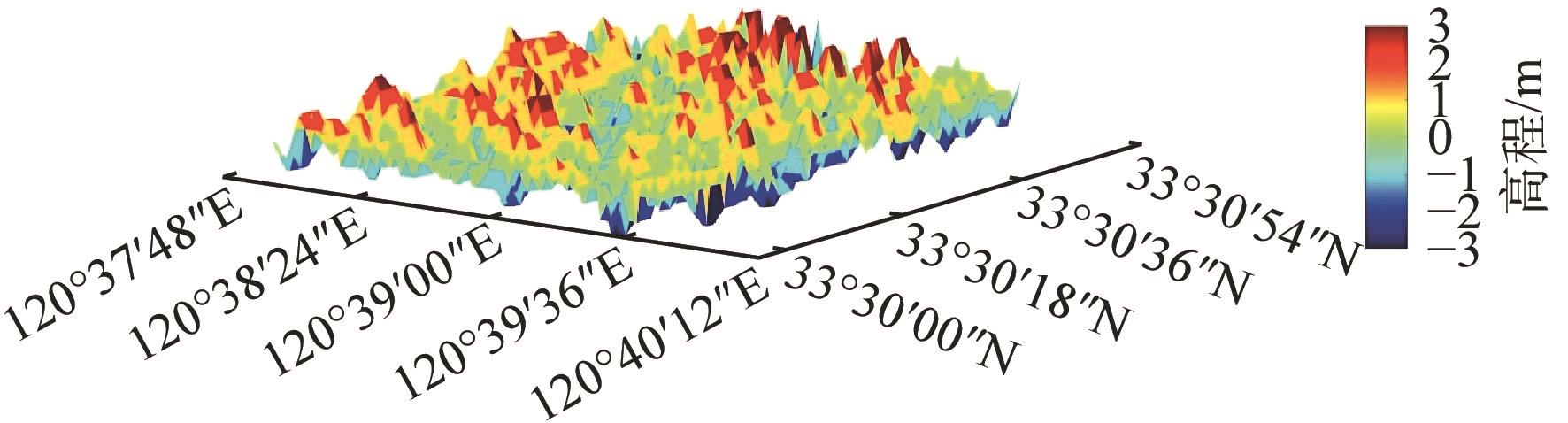
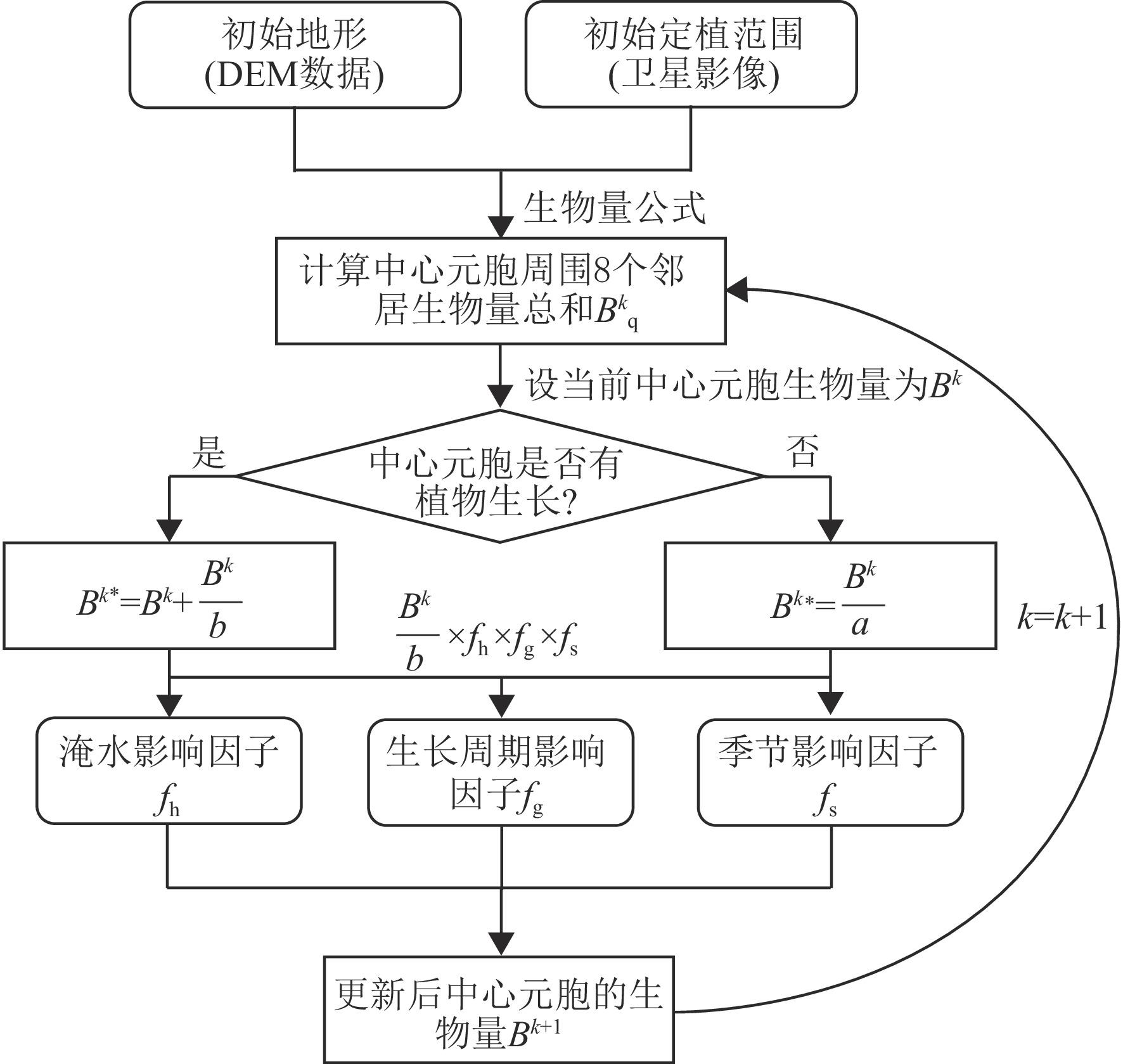
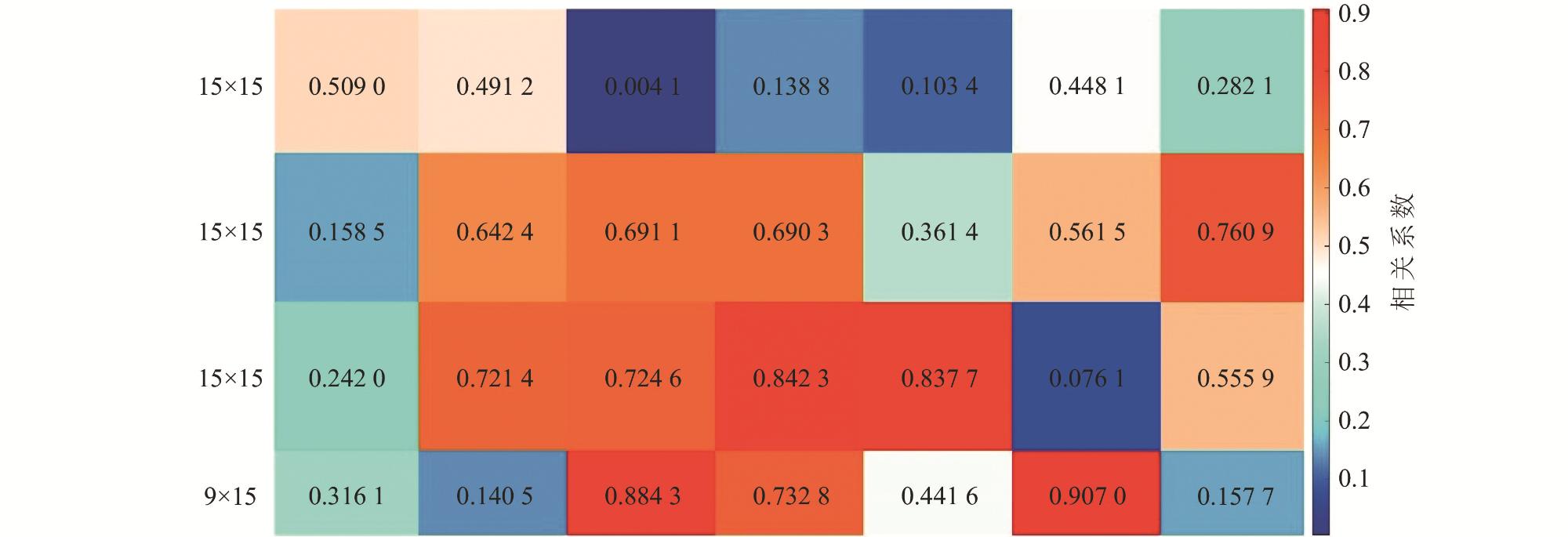
互花米草(Spartina alterniflora)入侵严重威胁滨海湿地生物多样性与生态功能,研究互花米草生长扩张过程对湿地生态和资源综合管理至关重要。本文构建了基于生物量概念的元胞自动机模型以模拟互花米草的时空分布,通过在计算中引入生物量,突破了传统模型仅能反映植被“有”或“无”状态的局限。模型采用摩尔邻域法则进行空间迭代计算,并综合考虑了淹水深度(表征水动力胁迫)、生长大周期(反映个体发育)、季节性物候变化等环境与生理约束。将这些因素作为修正系数动态调整元胞生长,以准确模拟盐沼生态系统中植被面积和密度变化的时空分布。选取江苏盐城湿地珍禽国家级自然保护区,以2000年5月的互花米草面积和密度分布为初始条件,对其15 a的生长扩张过程进行模拟。结果显示,2015年10月互花米草的密度分布模拟值与遥感解译数据之间的Pearson相关系数达到
蔡晗 , 李薇 . 基于生物量概念的互花米草时空分布元胞自动机模型[J]. 湿地科学, 2026 , 24(1) : 11 -21 . DOI: 10.13248/j.cnki.wetlandsci.20240226
The invasion of Spartina alterniflora (Smooth Cordgrass) in coastal wetlands has emerged as a pressing ecological challenge, significantly threatening native biodiversity and impairing critical ecosystem functions such as sediment stabilization, carbon sequestration, and habitat provision for migratory species. In response, comprehensive investigations into the spatiotemporal dynamics of this invasive species are urgently needed to inform effective ecosystem management and resource conservation strategies. This study addresses this imperative by developing an innovative biomass-based cellular automaton (CA) model specifically tailored to simulate the complex expansion patterns of S. alterniflora in salt marsh environments. Traditional CA models for vegetation dynamics often suffer from oversimplification, typically reducing vegetation presence to binary ‘on/off’ states without accounting for growth intensity or physiological responses. Our model advances this framework by incorporating biomass as a continuous variable, enabling nuanced representations of vegetation health, growth stages, and stress responses. The model employs Moore’s neighborhood rules for spatial iteration, which consider eight surrounding cells to more realistically capture seed dispersal and vegetative spread patterns. Key environmental and physiological constraints are integrated as dynamic correction factors: water depth (quantifying hydrodynamic stress), growth macrocycle (mapping individual developmental stages), and seasonal phenological changes (accounting for dormancy and active growth periods). This multi-parameter approach allows for accurate simulation of vegetation area and density distribution across heterogeneous wetland landscapes. The model was applied to the Jiangsu Yancheng Wetland National Nature Reserve, a representative coastal ecosystem, using May 2000 distribution data as initial conditions to simulate a 15-year expansion period. Validation against October 2015 remote sensing data demonstrated exceptional performance, with a Pearson correlation coefficient of
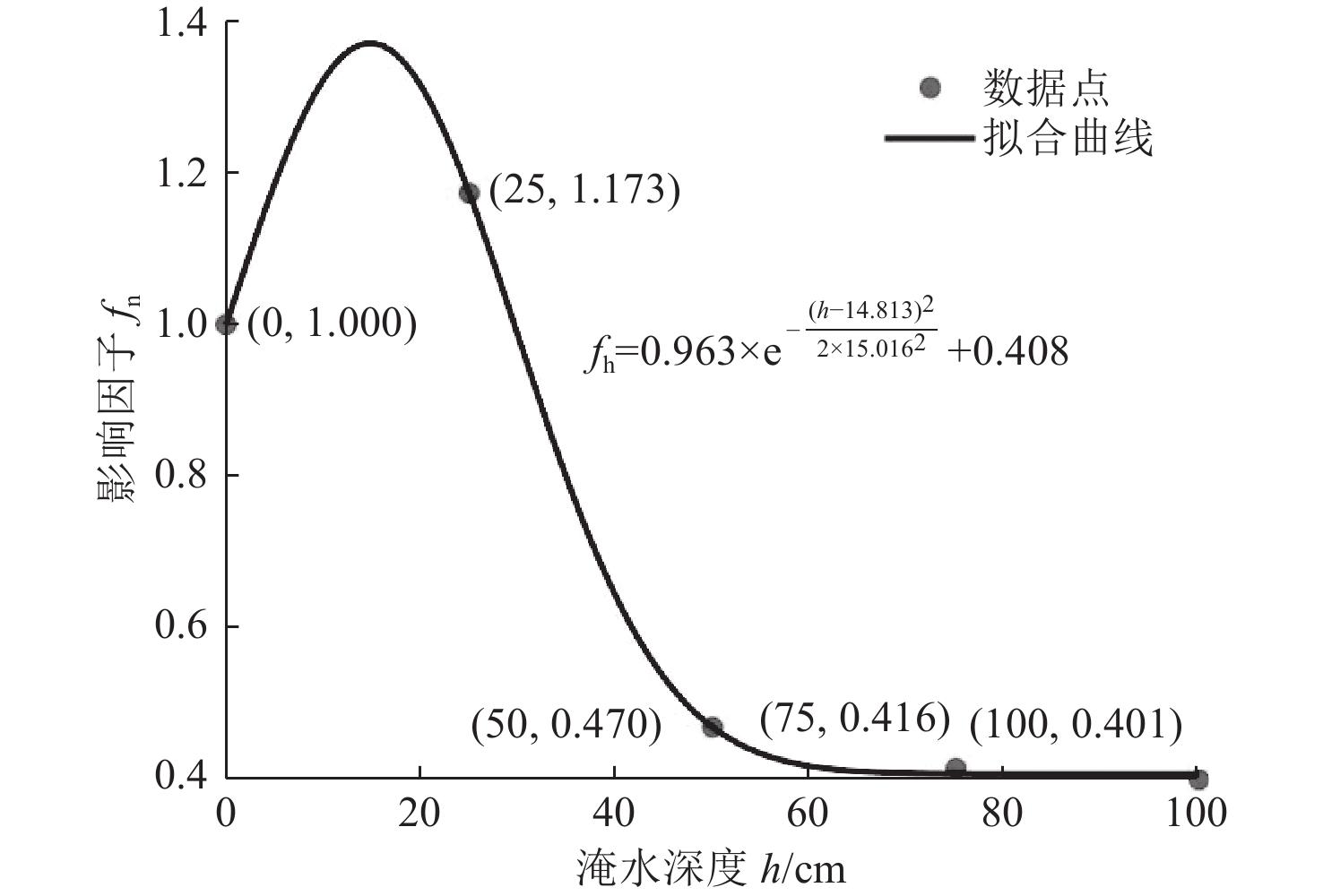
6 Curve graph of flooding impact factors corresponding to different flooding depths互花米草不同淹水深度对应的淹水影响因子曲线 |
2 Inundation impact factors corresponding to different water depths不同淹水深度对应的淹水影响因子 |
| 淹水深度 | 淹水影响因子 |
| 0 | 1 |
| 25 | 1.173 |
| 50 | 0.470 |
| 75 | 0.416 |
| 100 | 0.401 |
3 Growth cycle factors corresponding to different growth stages of Spartina alterniflora不同互花米草生长阶段对应的生长大周期因子 |
| 生长阶段 | 生物量累积比 | 生长大周期影响因子 |
| 生长初期 | 0~1/4 | 1 |
| 快速生长期 | 1/4~3/4 | c |
| 生长末期 | 3/4~1 | 1 |
4 Absolute growth rate (AGR) of aboveground biomass of Spartina alterniflora in different seasons[40]不同季节互花米草地上生物量的绝对增长率(AGR)[40] |
| 季节 | AGR/[g/(m2·a)] |
| 春 | 17.407 |
| 夏 | 10.800 |
| 秋 | −11.558 |
| 冬 | −2.546 |
5 Influencing factors of Spartina alterniflora corresponding to different seasons互花米草不同季节对应的影响因子 |
| 季节 | 季节影响因子 |
| 春 | 1.612 |
| 夏 | 1 |
| 秋 | −1.070 |
| 冬 | −0.236 |
7 Comparison of Spartina alterniflora plant density between remote sensing interpretation and simulation results2015年10月互花米草植被密度分布遥感解译与模拟结果比较 |
6 Simulation correlation coefficients of the biomass-based cellular automaton model under different expansion thresholds ( |
| K=185时r值 | 模拟过程r最大值 | |
| 注: | ||
| 0 | 0.824 7 | 0.830 4 (K=184) |
| 50 | 0.824 0 | 0.829 7 (K=184) |
| 100 | 0.823 8 | 0.829 5 (K=184) |
| 200 | 0.823 0 | 0.828 7 (K=184) |
| 400 | 0.820 0 | 0.825 7 (K=184) |
| 600 | 0.814 5 | 0.820 3 (K=184) |
| 800 | 0.806 5 | 0.812 2 (K=184) |
| 1 000 | 0.788 7 | 0.794 2 (K=184) |
7 Simulation correlation coefficients of the biomass-based cellular automaton model under different expansion rate parameter (a/b) combinations基于生物量概念的元胞自动机模型在不同扩张速率参数(a/b)组合下的模拟相关系数 |
| 参数 | 取值 | K=185时r值 | 模拟过程r最大值 |
| 注:a和b为共同表征互花米草扩张速率的模型参数。 | |||
| a/b | 64 | 0.796 4 | 0.847 5 (K=172) |
| 128 | 0.817 3 | 0.833 7 (K=184) | |
| 256 | 0.824 7 | 0.830 4 (K=184) | |
| 512 | 0.765 5 | 0.768 0 (K=184) | |
8 Simulation correlation coefficients of the biomass-based cellular automaton model under different growth cycle factors (c)基于生物量概念的元胞自动机模型在不同生长大周期影响因子(c)下的模拟相关系数 |
| 参数 | 取值 | K=185时的r值 | 模拟过程中r的最大值 |
| c | 1.0 | 0.823 0 | 0.828 1(K=184) |
| 1.2 | 0.824 2 | 0.829 6 (K=184) | |
| 1.3 | 0.824 6 | 0.830 1 (K=184) | |
| 1.4 | 0.824 7 | 0.830 2 (K=184) | |
| 1.5 | 0.824 7 | 0.830 4 (K=184) | |
| 1.6 | 0.824 7 | 0.830 4 (K=184) | |
| 1.7 | 0.824 3 | 0.830 1 (K=184) | |
| 1.8 | 0.824 1 | 0.829 9 (K=184) | |
| 2.0 | 0.823 5 | 0.829 3 (K=184) | |
| 3.0 | 0.822 0 | 0.828 5 (K=184) |
/
| 〈 |
|
〉 |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}