
|
樊龙凤(1993—),女,甘肃省泾川人,博士研究生,主要从事滨海湿地碳循环方面的研究。E-mail: fanlongflying@126.com |
收稿日期: 2024-09-18
修回日期: 2024-10-26
网络出版日期: 2026-03-12
版权
Carbon sequestration potential of coastal salt marshes in Yancheng under the invasion of Spartina alterniflora
Received date: 2024-09-18
Revised date: 2024-10-26
Online published: 2026-03-12
Copyright
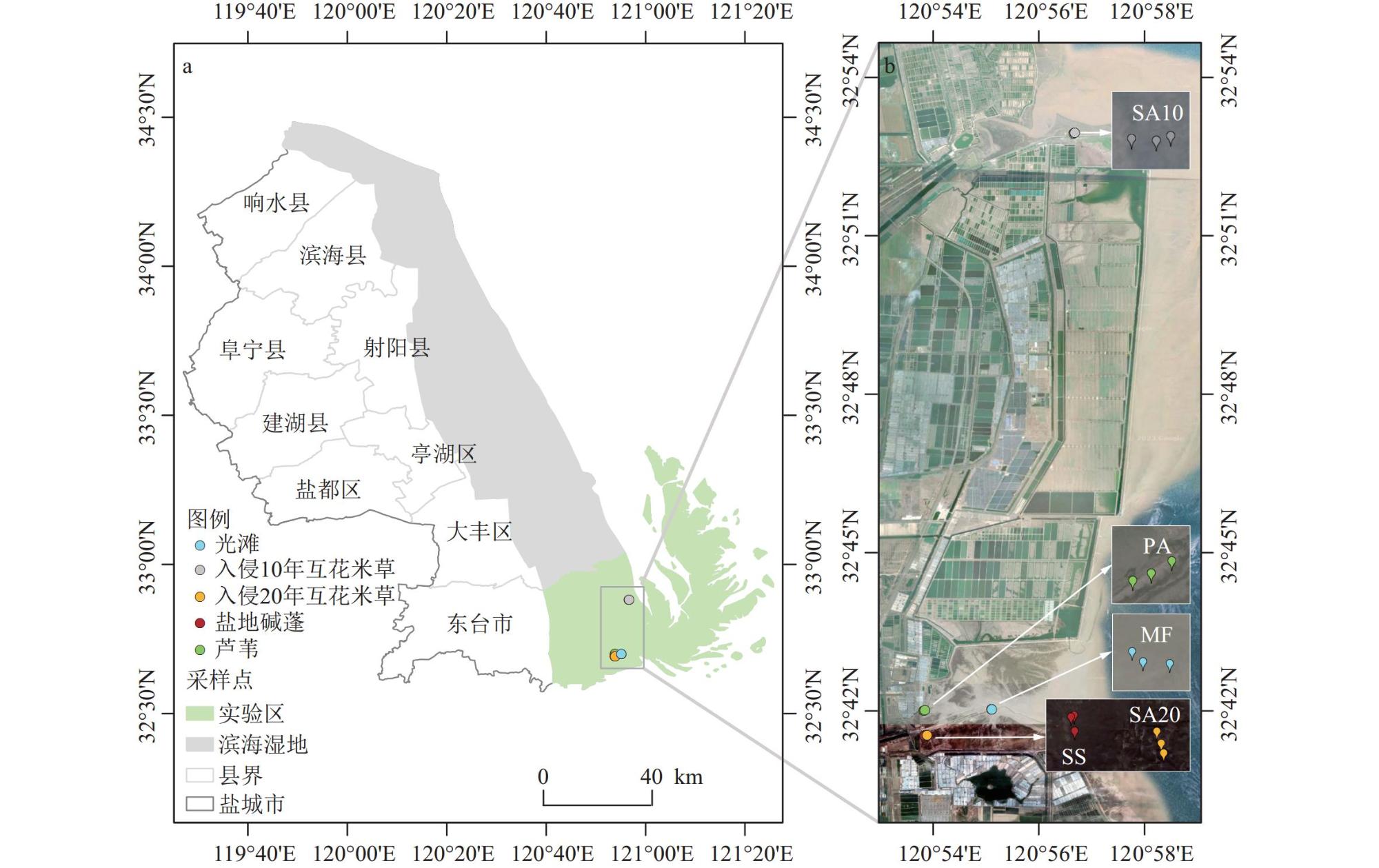
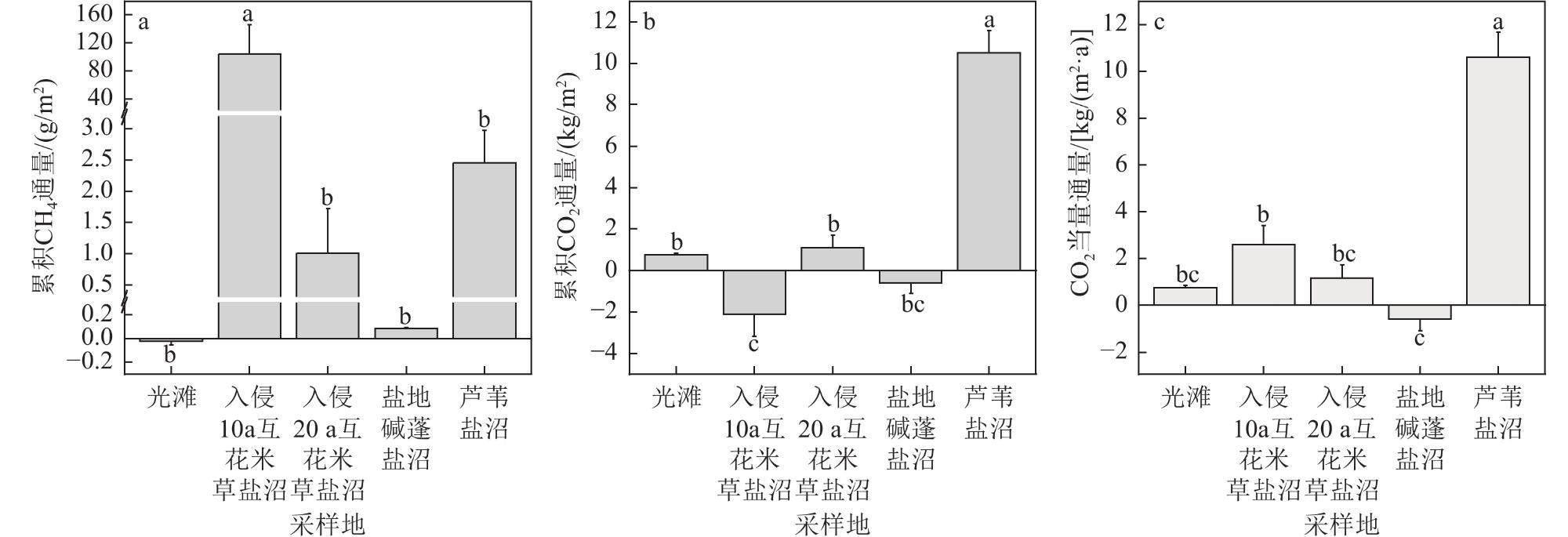
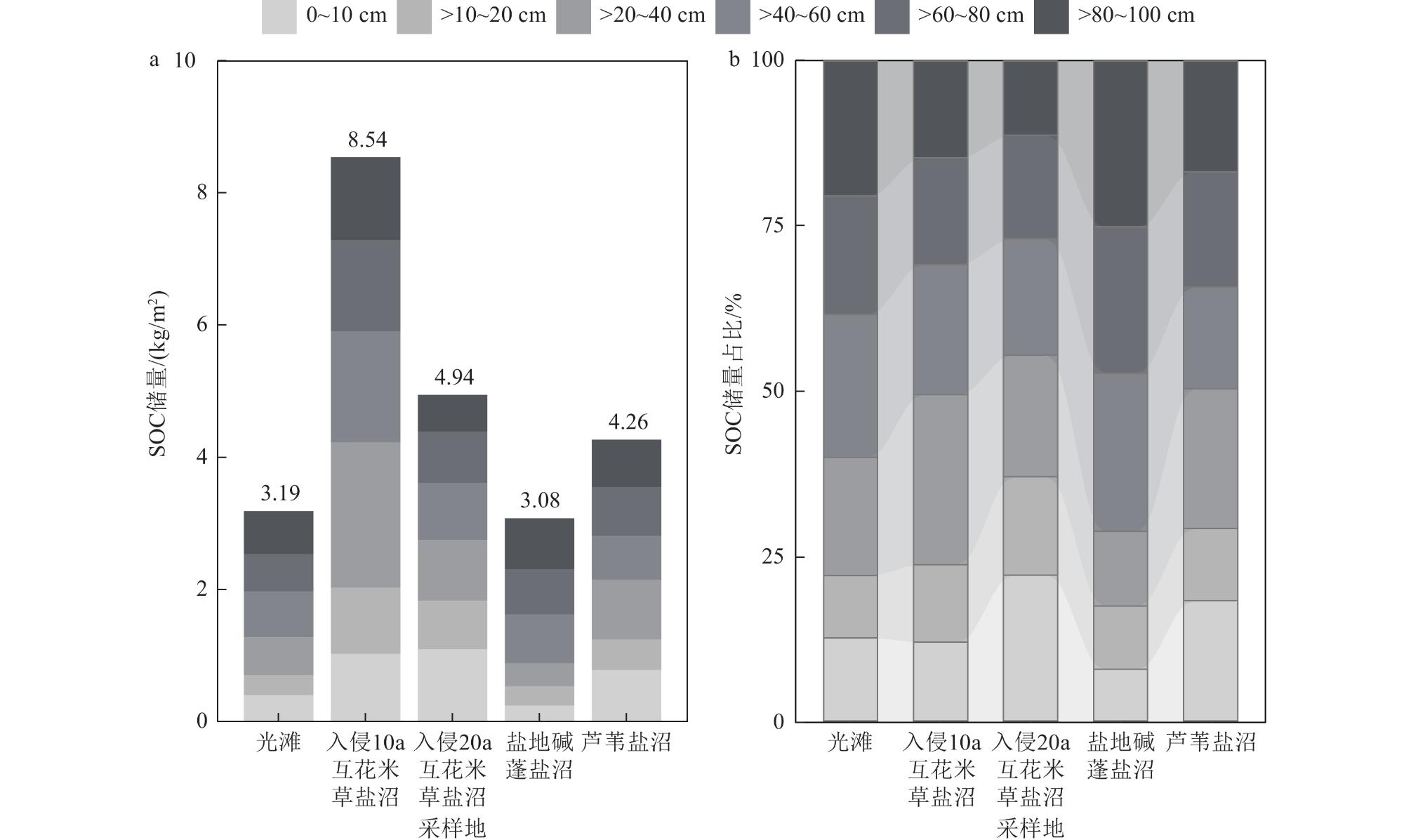
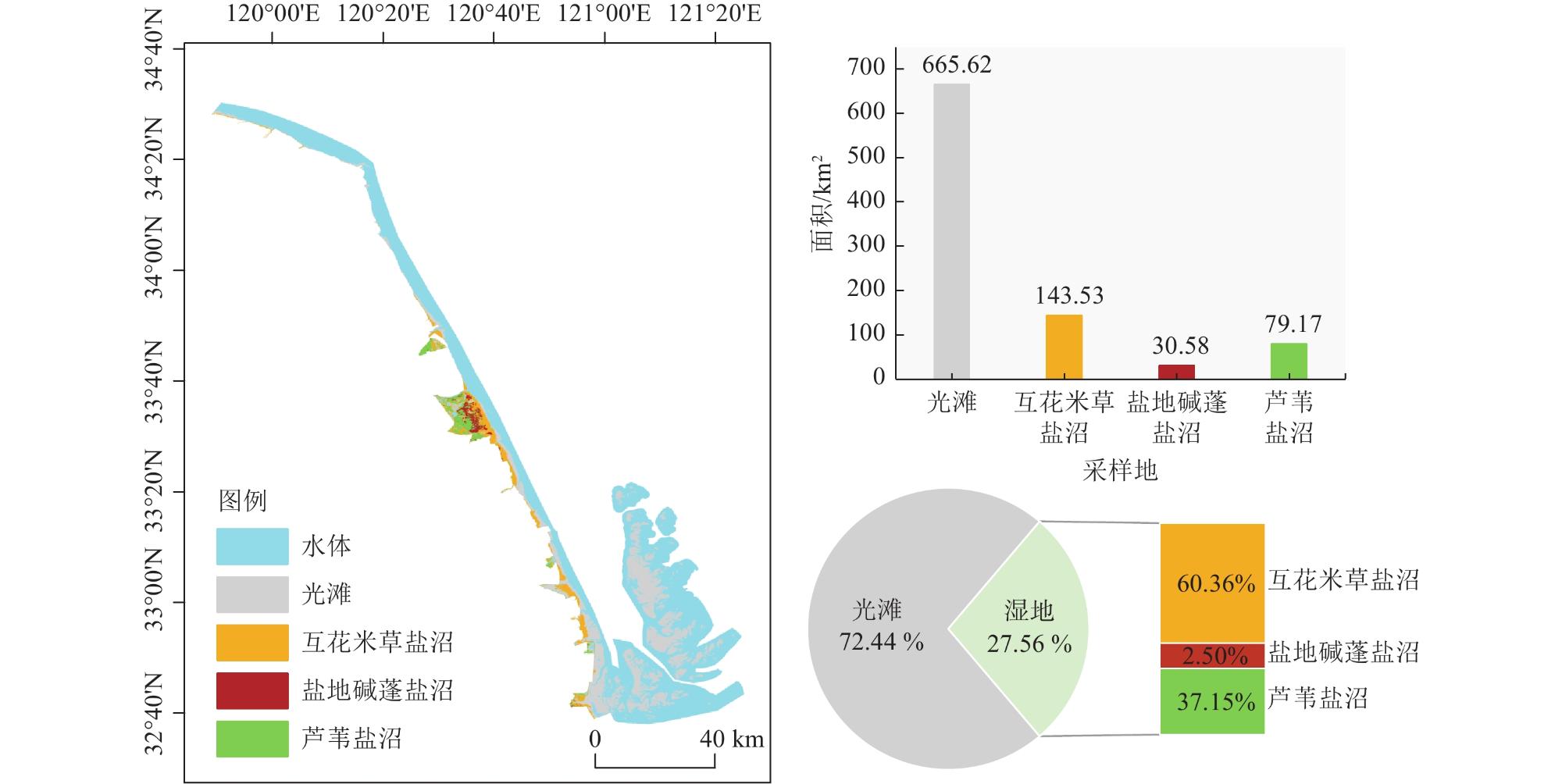
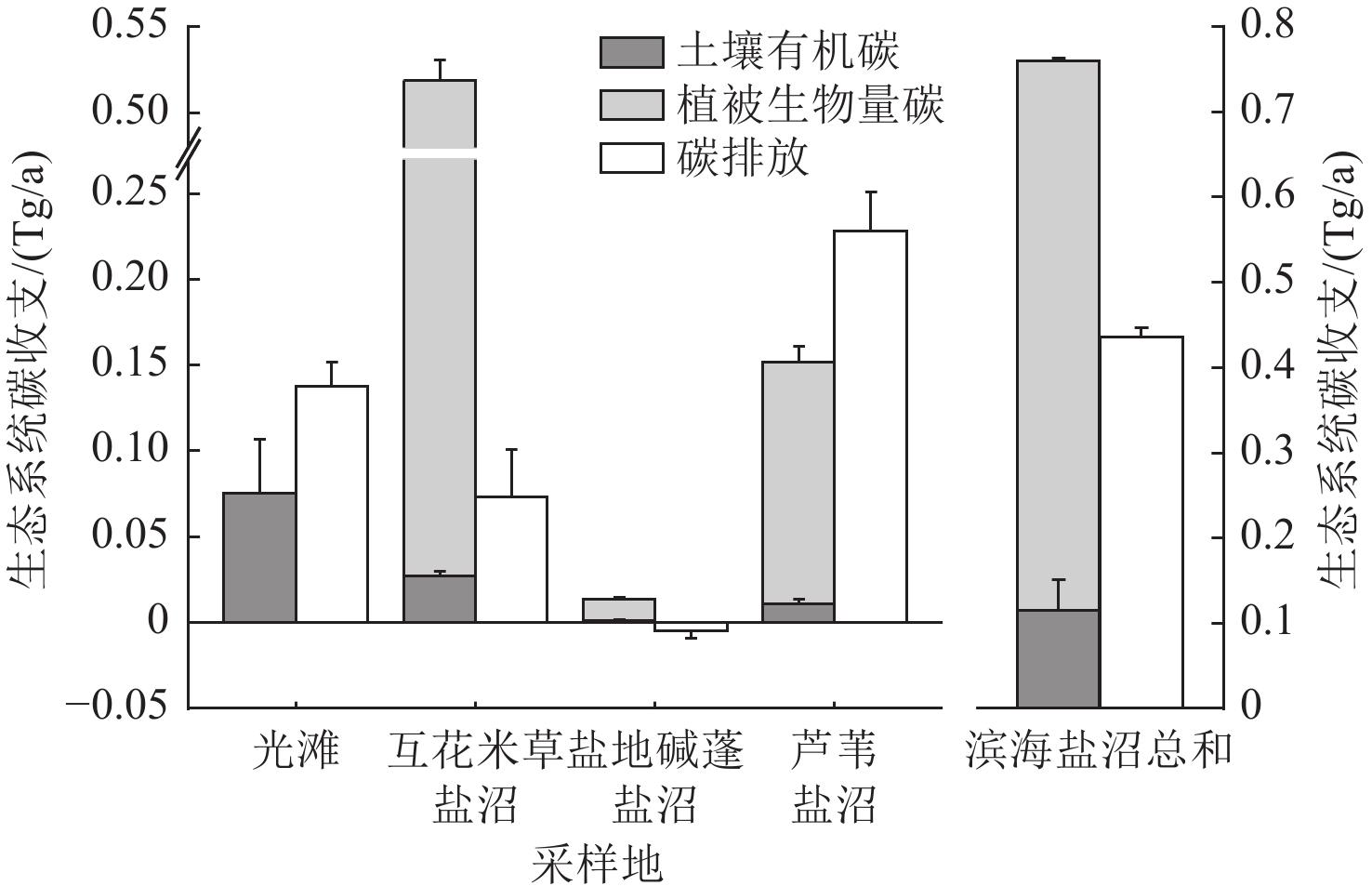
海岸带蓝碳生态系统正面临互花米草(Spartina alterniflora)入侵的威胁。为了全面评估互花米草入侵对盐城市滨海盐沼碳汇潜力的影响,对盐城市典型盐沼,包括入侵10 a和20 a的互花米草盐沼以及本土光滩、盐地碱蓬(Suaeda salsa)盐沼和芦苇(Phragmites australis)盐沼进行了碳通量(CH4和CO2)、植被生物量和土壤有机碳含量的测定和分析。研究结果表明,互花米草盐沼的碳排放总量低于芦苇盐沼,但高于光滩和盐地碱蓬盐沼。相比本土盐沼,互花米草入侵增加了植被和土壤碳库的储量。盐城市滨海盐沼每年的碳排放量(0.44 Tg/a)低于碳输入量(0.76 Tg/a),归因于互花米草群落扩张带来的碳固定和盐地碱蓬盐沼的负排放。2021年,盐城市滨海盐沼的碳汇潜力为3.73 Tg,较1987年减少了51.6%。其中,互花米草入侵增加的碳汇(1.39 Tg)未能补偿盐地碱蓬和芦苇盐沼面积萎缩带来的碳汇损失(1.59 Tg)。除湿地面积萎缩直接导致的土壤碳库损失外,互花米草长期入侵所引发的湿地环境变化也会间接削弱区域碳汇功能。
樊龙凤 , 程俊翔 , 徐力刚 , 蒋名亮 , 许加星 . 互花米草入侵下盐城滨海盐沼碳汇潜力研究[J]. 湿地科学, 2025 , 23(6) : 1277 -1287 . DOI: 10.13248/j.cnki.wetlandsci.20240261
Coastal wetlands play a crucial role in mitigating climate change due to their “blue carbon” function. However, the carbon sequestration capacity of these ecosystems is threatened by the invasion of Spartina alterniflora. Salt marshes represent the largest type of coastal wetland in China. To comprehensively assess the impact of Spartina alterniflora invasion on the carbon sequestration capacity of salt marsh ecosystems, we conducted measurements and analyses of carbon flux (e.g., methane [CH4] and carbon dioxide [CO2]), vegetation biomass, and soil organic carbon (SOC) content in typical salt marshes (including marshes invaded by exotic Spartina alterniflora for 10 a and 20 a, as well as native mudflat, Suaeda salsa marsh and Phragmites australis marsh) in Yancheng and assessed the carbon sequestration potential at the regional scale. The study results revealed that the total carbon emissions (comprising CO2 and CH4 fluxes) from Spartina alterniflora salt marshes (1.16-2.58 kg/(m2·a)) were lower than those from Phragmites australis salt marshes (10.60 kg/(m2·a)) but higher than those from bare mudflats and Suaeda salsa salt marshes (0.75 and −0.60 kg/(m2·a), respectively). Compared to the native salt marshes, the invasion of Spartina alterniflora led to an increase in the storage of both vegetation and soil carbon pools. Specifically, the high primary productivity of Spartina alterniflora significantly enhanced biomass carbon accumulation above and below ground. Concurrently, SOC content in the invaded marshes showed a notable increase compared to the native mudflat and Suaeda salsa marsh, particularly in the upper soil layers, although the magnitude of this increase varied with the duration of invasion. From an ecosystem carbon budget perspective, the annual carbon emissions from Yancheng coastal salt marshes (0.44 Tg/a) were lower than the total annual carbon input (0.76 Tg/a), attributed to carbon fixation associated with the expansion of Spartina alterniflora and the negative emissions from Suaeda salsa salt marshes. Despite the current net sink status, a historical analysis of carbon stocks revealed a significant decline in the overall carbon sequestration potential of the Yancheng coastal salt marshes. By 2021, the total estimated carbon sequestration potential was 3.73 Tg. This represents a substantial reduction of 51.6% compared to the estimated potential in 1987. This dramatic decrease is primarily driven by large-scale losses in the areal extent of native salt marshes due to land reclamation, coastal development, and ecological degradation. Crucially, the increase in carbon sequestration resulting from the areal expansion of Spartina alterniflora (contributing an estimated +1.39 Tg) was insufficient to offset the massive carbon sink loss caused by the shrinkage of native Suaeda salsa and Phragmites australis salt marshes (estimated at -1.59 Tg collectively). In addition to the direct loss of soil carbon stocks resulting from wetland area shrinkage due to anthropogenic reclamation, the long-term environmental changes in wetlands under Spartina alterniflora invasion also indirectly weakened the regional carbon sequestration function. The findings provide crucial scientific evidence and foundational data support for the management of carbon sequestration resources and the conservation of coastal salt marsh ecosystems.
1 Carbon content in plant biomass of the coastal salt marshes in Yancheng City /(kg/m2)盐城市滨海盐沼植物生物量碳含量/(kg/m2) |
| 器官 | 入侵10 a互花米草盐沼 | 入侵20 a互花米草盐沼 | 盐地碱蓬盐沼 | 芦苇盐沼 |
| 注:表中数据为(平均值±标准误差)。数据上标小写字母不同表示不同盐沼之间差异显著(p<0.05)。 | ||||
| 地上活体植物 | (1.01±0.08)a | (0.57±0.05)ab | (0.22±0.01)b | (0.59±0.03)ab |
| 立枯物 | (0.28±0.01)a | (0.32±0.06)a | (0.06±0.00)b | (0.29±0.00)a |
| 地下1 m内根系 | (3.07±0.24)a | (1.56±0.15)b | (0.16±0.01)c | (0.93±0.07)b |
| 总生物量 | (4.36±0.33)a | (2.39±0.20)b | (0.41±0.01)c | (1.78±0.12)bc |
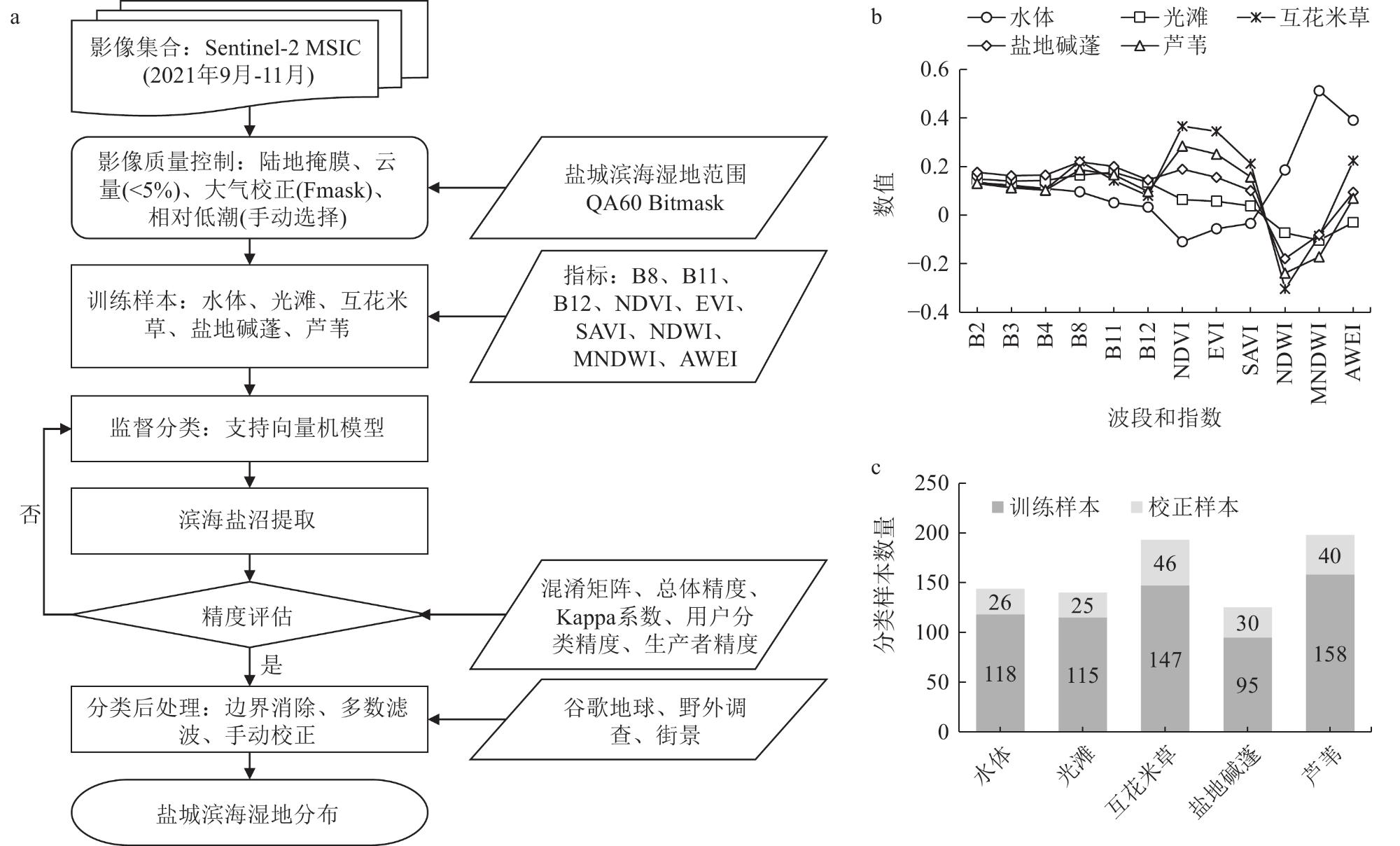
2 Classification of coastal salt marshes in Yancheng City based on Support Vector Machine基于支持向量机的盐城市滨海盐沼分类结果 |
| 总体分类 精度 | 水体 | 光滩 | 互花米草 盐沼 | 盐地碱蓬 盐沼 | 芦苇 盐沼 | 用户分类 精度/% |
| 注:Kappa系数为0.969 7。 | ||||||
| 水体 | 26 | 0 | 0 | 0 | 0 | 96.3 |
| 光滩 | 1 | 24 | 0 | 0 | 0 | 92.3 |
| 互花米草盐沼 | 0 | 2 | 44 | 0 | 0 | 100.0 |
| 盐地碱蓬盐沼 | 0 | 0 | 0 | 30 | 0 | 96.8 |
| 芦苇盐沼 | 0 | 0 | 0 | 1 | 39 | 100.0 |
| 生产者精度/% | 100.0 | 96.0 | 95.7 | 100.0 | 97.5 | 97.6 |
3 Variation of carbon sink potential in the coastal salt marshes of Yancheng City from 1987 to 20211987—2021年盐城市滨海盐沼碳汇潜力变化 |
| 采样地 | 湿地面积/km2 | 碳排放/(Tg/a) | 植被碳库/Tg | 土壤碳库/Tg | 碳汇潜力/Tg | |||||||||
| 1987 | 2021 | 1987 | 2021 | 1987 | 2021 | 1987 | 2021 | 1987 | 2021 | |||||
| 注: 1987年的盐沼面积引自左平等[34]。互花米草的数据为入侵10 a和20 a互花米草盐沼数据的平均值。 | ||||||||||||||
| 光滩 | 1 926.09 | 665.62 | 0.40 | 0.140 | 6.140 | 2.12 | 5.74 | 1.98 | ||||||
| 互花米草盐沼 | 0.49 | 143.53 | −0.00 | 0.070 | 0.002 | 0.49 | 0.003 | 0.97 | 0.01 | 1.39 | ||||
| 盐地碱蓬盐沼 | 411.55 | 30.58 | −0.07 | −0.005 | 0.170 | 0.01 | 1.270 | 0.09 | 1.50 | 0.11 | ||||
| 芦苇盐沼 | 143.94 | 79.17 | 0.42 | 0.230 | 0.260 | 0.14 | 0.610 | 0.34 | 0.45 | 0.25 | ||||
| 合计 | 2 482.07 | 918.90 | 0.75 | 0.440 | 0.430 | 0.64 | 8.020 | 3.52 | 7.70 | 3.73 | ||||
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}