
中国南方滨海湿地土壤氧化亚氮还原菌丰度对互花米草入侵的响应及其关键影响因素
|
叶悦龄(2003—),女,福建省漳州人,本科生,从事滨海湿地氮循环研究。E-mail: 2095408116@qq.com |
收稿日期: 2024-08-20
修回日期: 2024-10-05
网络出版日期: 2026-03-12
版权
Response of soil nitrous oxide reducing bacteria abundance to Spartina alterniflora invasion in coastal wetlands of southern China and its key influencing factors
Received date: 2024-08-20
Revised date: 2024-10-05
Online published: 2026-03-12
Copyright
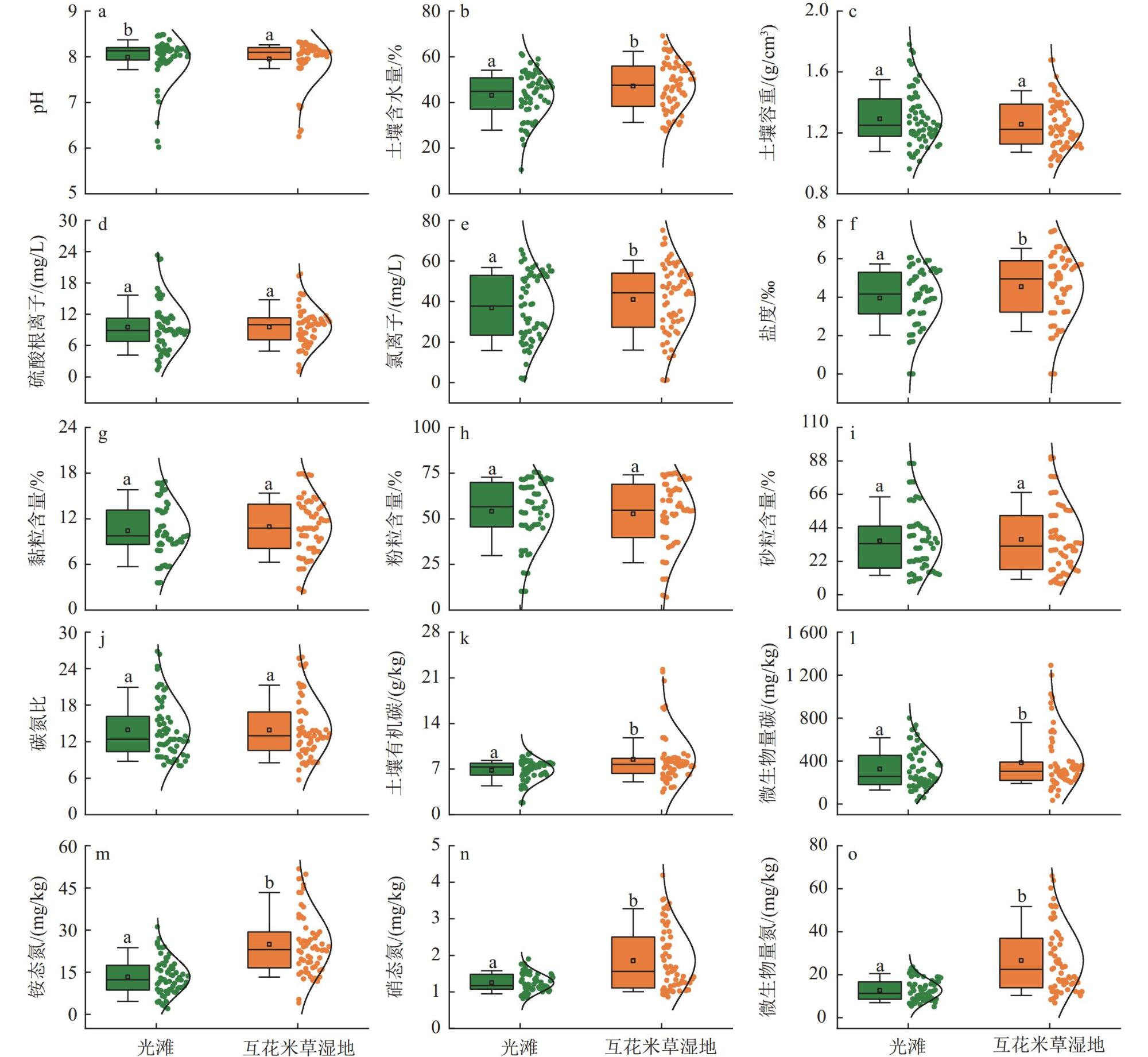
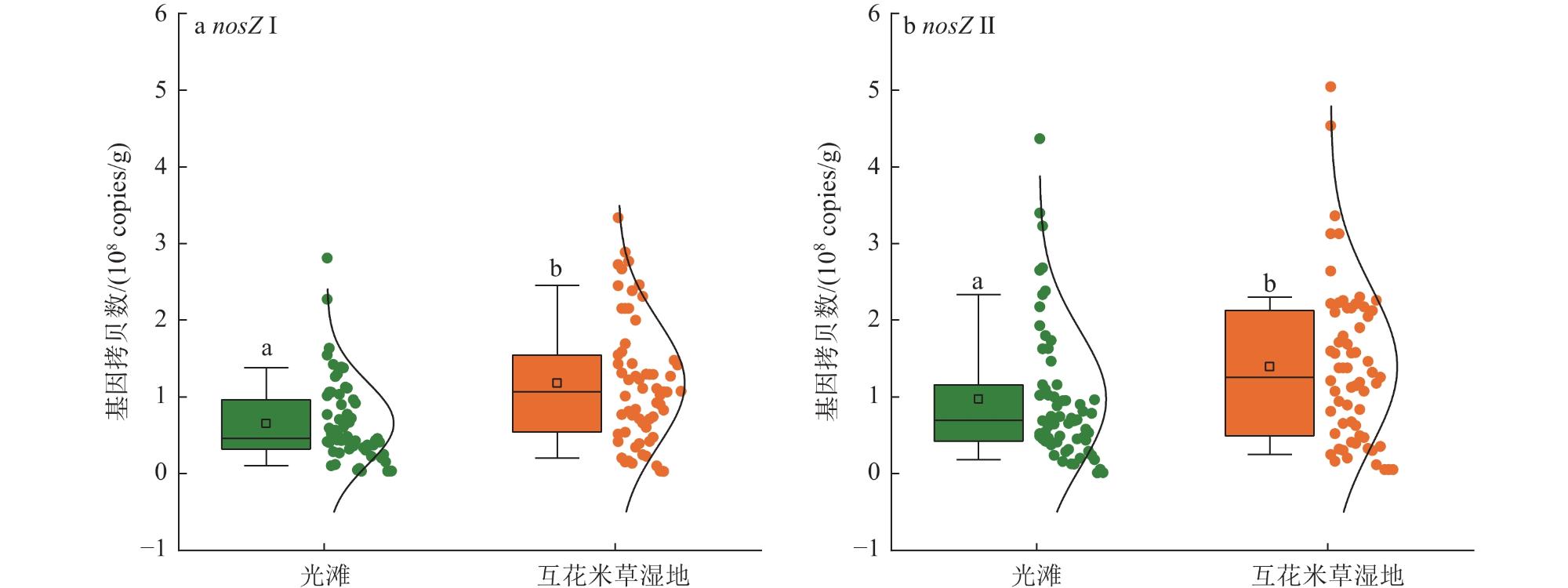
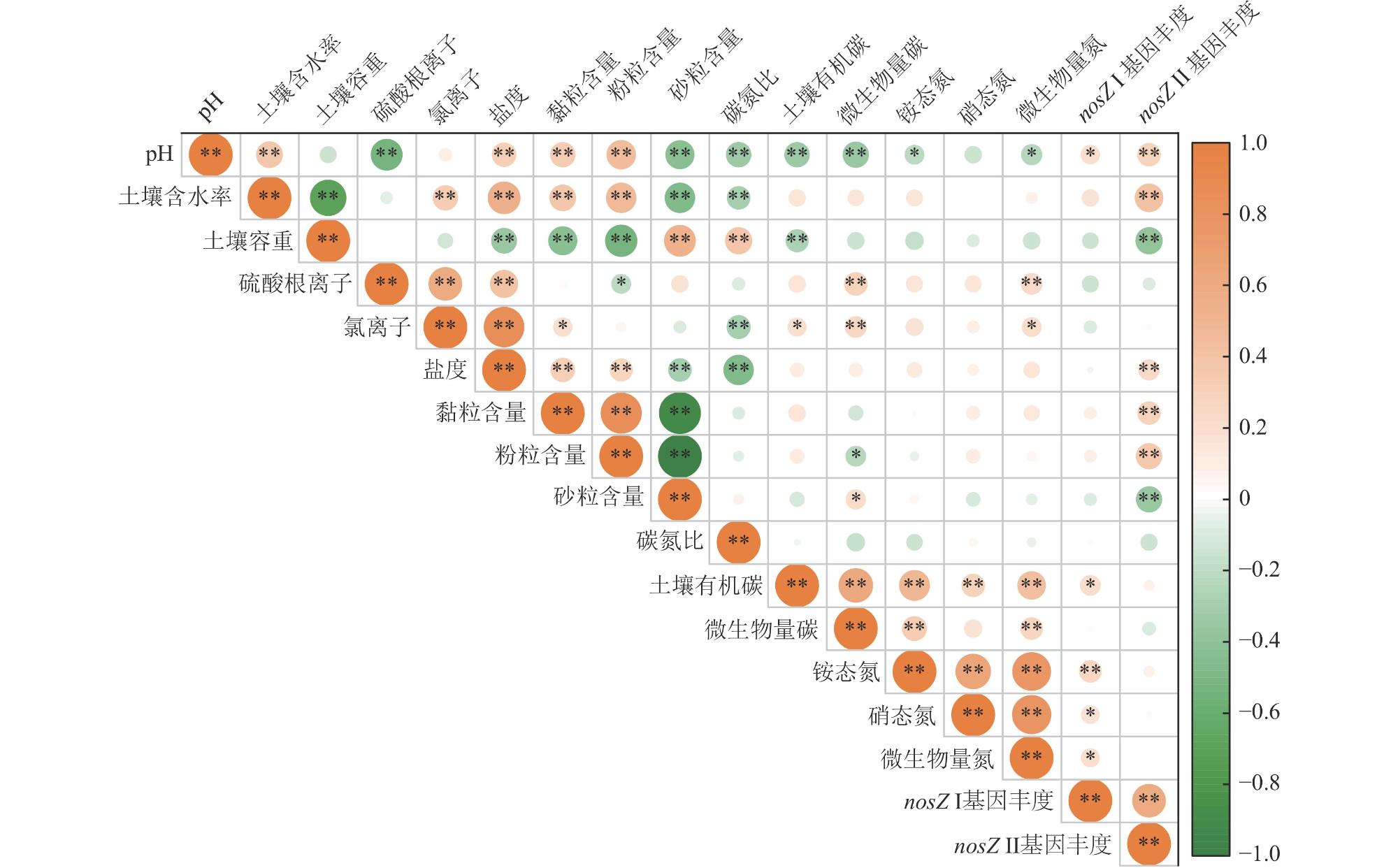
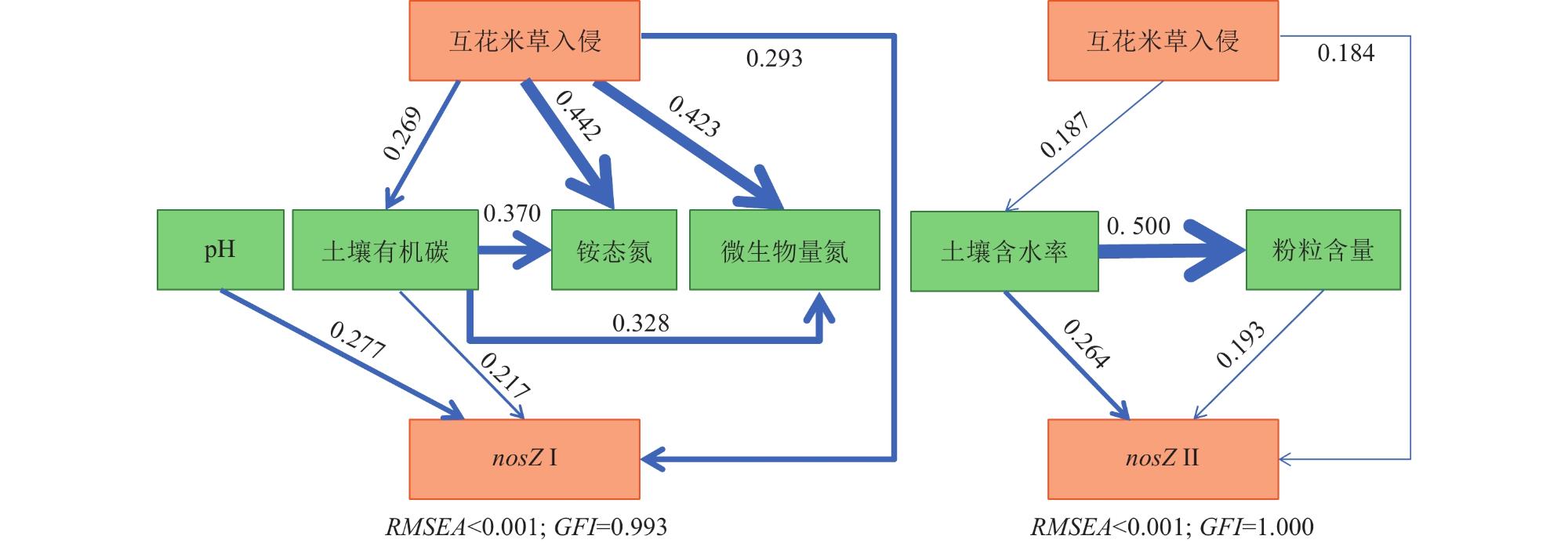
为揭示滨海湿地土壤氧化亚氮还原菌丰度对互花米草(Spartina alterniflora)入侵的响应及其关键影响因素,在中国南方21处滨海湿地采集土壤样品,采用实时荧光定量PCR技术,测定光滩和互花米草湿地土壤氧化亚氮还原菌功能基因丰度。结果表明,光滩和互花米草湿地土壤nosZ Ⅱ基因拷贝数显著高于nosZ Ⅰ,表明nosZ Ⅱ型氧化亚氮还原菌在中国南方滨海湿地中占主导地位。互花米草湿地土壤nosZ Ⅰ和nosZ Ⅱ基因拷贝数均显著高于光滩,表明互花米草入侵显著提高了滨海湿地土壤氧化亚氮还原菌丰度。逐步回归分析表明,土壤pH和含水率分别是nosZ Ⅰ和nosZ Ⅱ基因丰度的最重要调控因子。土壤nosZ I基因丰度与土壤pH、有机碳、铵态氮、硝态氮和微生物生物量氮呈显著正相关;而nosZ Ⅱ基因丰度与土壤pH、含水率、盐度和黏粒含量呈显著正相关,与土壤容重和沙粒含量呈显著负相关。结构方程模型分析表明,互花米草入侵可直接影响nosZ I和nosZ Ⅱ基因丰度,也可通过影响土壤有机碳含量和含水率分别间接影响nosZ I和nosZ Ⅱ基因丰度。综上所述,互花米草入侵显著提高了滨海湿地土壤氧化亚氮还原菌丰度,土壤pH和含水率分别是调控nosZ Ⅰ和nosZ Ⅱ基因丰度的关键环境因子。
叶悦龄 , 朱思思 , 左婷 , 杨平 , 靳少非 , 魏翔莺 , 林永新 , 叶桂萍 . 中国南方滨海湿地土壤氧化亚氮还原菌丰度对互花米草入侵的响应及其关键影响因素[J]. 湿地科学, 2025 , 23(4) : 803 -812 . DOI: 10.13248/j.cnki.wetlandsci.20240233
To investigate the response of soil nitrous oxide (N2O) reducing bacterial communities to Spartina alterniflora invasion and to identify the key environmental drivers shaping these responses, we conducted a large-scale field survey across 21 coastal wetlands in southern China. Soil samples were collected from both bare tidal flats and adjacent S. alterniflora invaded sites. Using quantitative real-time PCR, we quantified the abundance of nosZ I and nosZ II genes, which encode distinct clades of nitrous oxide reductase enzymes responsible for the microbial reduction of N2O to dinitrogen (N2), the final step of the denitrification process. Our results revealed that the copy number of the nosZ II gene was significantly higher than that of nosZ I in both vegetation types, suggesting that nosZ II-type N2O reducers dominate the microbial N2O sink in subtropical coastal wetland soils. Notably, both nosZ I and nosZ II gene abundances were significantly elevated in soils invaded by S. alterniflora, indicating that plant invasion enhances the capacity of soil microbial communities to reduce N2O emissions. This finding is of particular ecological relevance, as coastal wetlands serve as both sources and sinks of N2O. Stepwise regression analyses identified soil pH and moisture content as the most influential factors controlling the abundance of nosZ I and nosZ II genes, respectively. Specifically, nosZ I gene abundance was positively correlated with pH, soil organic carbon, ammonium and nitrate nitrogen, and microbial biomass nitrogen. In contrast, nosZ II gene abundance showed significant positive relationships with pH, water content, salinity, and clay content, and negative correlations with soil bulk density and sand fraction. These contrasting patterns suggest that nosZ I and nosZ II organisms may occupy distinct ecological niches and respond differently to environmental change. Furthermore, structural equation modeling indicated that S. alterniflora invasion influences nosZ gene abundances through both direct and indirect pathways. While invasion directly stimulated the abundance of both gene clades, it also indirectly influenced them by modifying key soil properties, such as increasing organic carbon availability and altering water retention. Taken together, our study highlights that S. alterniflora invasion enhances the microbial potential for N2O reduction in coastal wetlands and that soil pH and moisture are critical environmental filters shaping the structure and function of N2O-reducing communities. These findings underscore the importance of considering plant invasion effects when evaluating greenhouse gas dynamics in coastal ecosystems and suggest that managing vegetation transitions may represent a viable strategy to enhance the soil microbial sink capacity for N2O under global change scenarios.
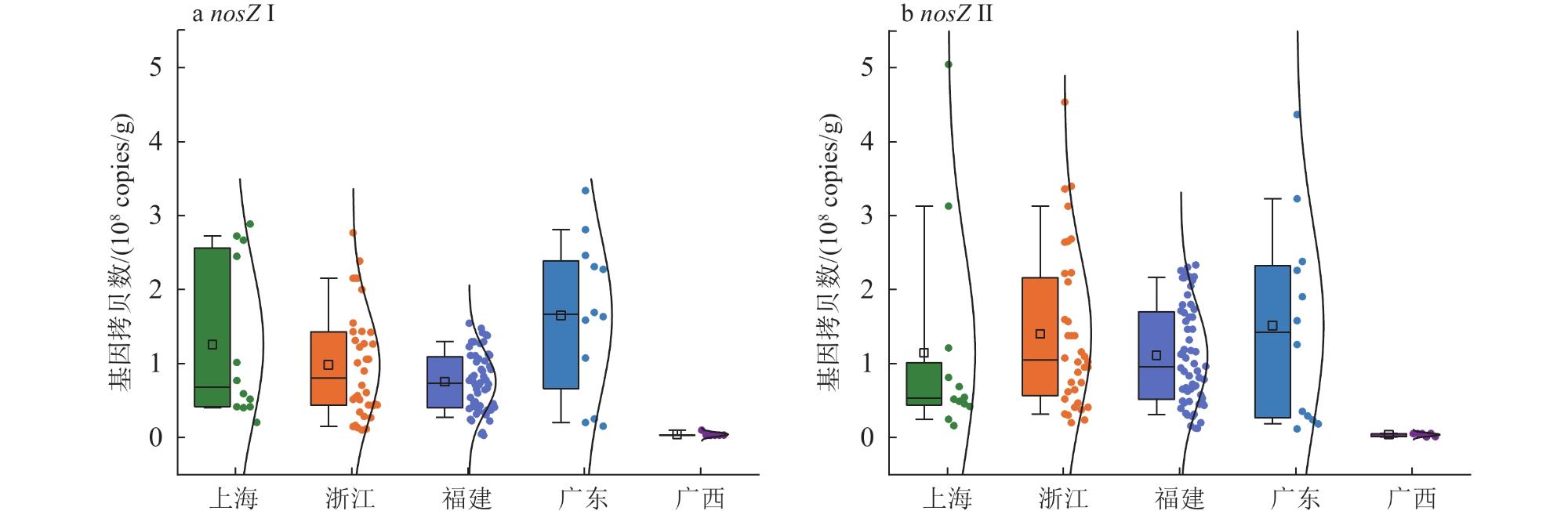
1 Distribution of soil sampling sites in coastal wetlands of southern China中国南方滨海湿地土壤采样点分布 |
| 样点 | 经纬度 |
| 上海崇明区 | 31°29'28"N,121°52'04"E |
| 上海奉贤区 | 30°50'39"N,121°37'23"E |
| 浙江绍兴 | 30°12'23"N,120°47'16"E |
| 浙江舟山 | 29°59'10"N,122°13'10"E |
| 浙江台州 | 29°09'15"N,121°34'14"E |
| 浙江温岭 | 28°18'39"N,121°34'07"E |
| 浙江温州 | 27°48'10"N,120°48'00"E |
| 浙江温州 | 27°35'22"N,120°38'04"E |
| 福建福鼎 | 27°06'49"N,120°16'19"E |
| 福建宁德 | 26°58'18"N,120°10'24"E |
| 福建福州 | 26°01'34"N,119°38'05"E |
| 福建福清 | 25°29'14"N,119°14'21"E |
| 福建福清 | 25°37'49"N,119°28'33"E |
| 福建莆田 | 25°13'54"N,119°08'59"E |
| 福建莆田 | 25°24'19"N,119°07'47"E |
| 福建漳州 | 24°01'10"N,117°44'27"E |
| 福建漳州 | 24°11'08"N,117°57'25"E |
| 福建南安 | 24°40'53"N,118°26'12"E |
| 广东台山 | 21°56'34"N,112°46'00"E |
| 广东雷州 | 20°53'41"N,110°10'23"E |
| 广西北海 | 21°35'17"N,109°37'05"E |
4 Correlation of the abundance of nitrous oxide reducing bacteria and soil physicochemical properties in coastal wetlands in southern China中国南方滨海湿地土壤氧化亚氮还原菌功能基因丰度与土壤理化性质之间的相关关系 p < 0.05,**表示p < 0.01。]]> |
2 Stepwise regression analysis of environmental factors related to the abundance of soil nitrous oxide reducing bacteria in coastal wetlands of southern China中国南方滨海湿地与土壤氧化亚氮还原菌丰度相关的环境因子的逐步回归分析 |
| 基因 | 环境因子 | 标准化回归系数 | p |
| nosZ Ⅰ | pH | 0.344 | <0.001 |
| 土壤有机碳 | 0.254 | <0.05 | |
| 铵态氮 | 0.235 | <0.05 | |
| 氯离子 | −0.206 | <0.05 | |
| nosZ Ⅱ | 土壤含水率 | 0.317 | <0.001 |
| 粉粒含量 | 0.207 | <0.05 | |
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}