
水深梯度对苦草典型功能性状的影响
|
容鑫林(1996—),男,安徽省怀宁人,硕士研究生,从事生物资源保护利用研究。E-mail: 1396919031@qq.com |
收稿日期: 2023-02-19
修回日期: 2023-04-07
网络出版日期: 2026-03-12
版权
Impact of water depth gradient on the typical functional traits of Vallisneria natans
Received date: 2023-02-19
Revised date: 2023-04-07
Online published: 2026-03-12
Copyright
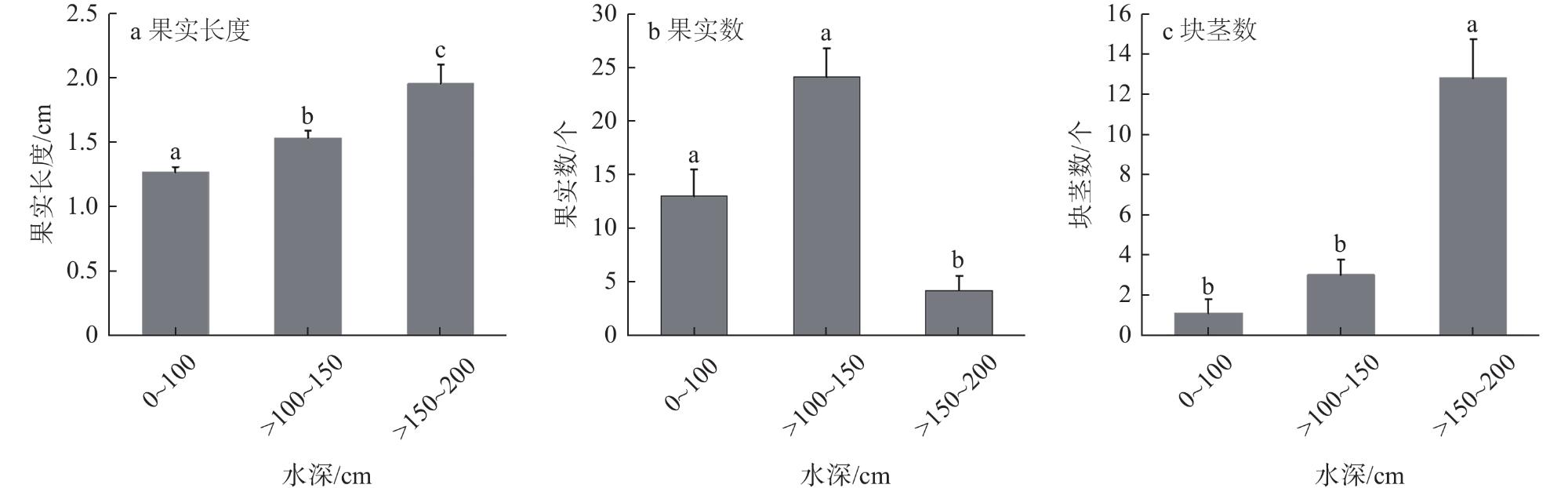
苦草(Vallisneria natans)是通江湖泊优势物种,对于维持湖泊生态系统的结构和功能具有重要作用。以安徽大学磬苑校区内人工景观湖琵琶湖生态修复后人工种植的苦草种群为研究对象,通过实地监测,了解苦草在生长周期的不同阶段(种子萌发、生长和繁殖等),其典型功能性状对3种不同水深条件的响应。研究结果显示,水深对苦草的典型功能性状具有影响,苦草种子的萌发和幼苗的存活率与水深呈负相关关系;随着水深的增加,苦草株高逐渐增加,叶片长度和比叶面积均提高,且地上生物量占比在水深最大条件下最大;此外,苦草植株在浅水区(0~100 cm、>100~150 cm水深)主要进行有性生殖,在深水区(>150~200 cm水深)主要通过地下块茎进行克隆繁殖。可以为水位波动条件下苦草种群的保护和恢复提供一定的理论支持。
容鑫林 , 周忠泽 , 王淑琼 . 水深梯度对苦草典型功能性状的影响[J]. 湿地科学, 2025 , 23(1) : 140 -149 . DOI: 10.13248/j.cnki.wetlandsci.20230040
Vallisneria natans is a dominant species in lakes connecting the Yangtze River, and is essential in maintaining the structure and function of lake ecosystems. Water depth is the main environmental factor affecting the survival and growth of submerged macrophytes. Too shallow or too deep water condition is not conducive to the stability of submerged macrophytes. In practice, due to the influence of human factors and differences in regional conditions, the water level of many aquatic ecosystems fluctuates to a certain extent, which may have a certain impact on the growth and reproduction of submerged macrophytes, for instance, Vallisneria natans. However, the specific influence of water depth on Vallisneria natans is still unclear. In this study, the population of Vallisneria natans planted after ecological restoration of Lake Pipa of Anhui University was selected as the research object. Three sampling areas with relatively consistent growth condition of Vallisneria natans were selected. In each sampling area, three sampling points were selected according to three different water depth, i.e., 0-10 cm, 10-50 cm, and 50-90 cm (at early stage), and 0-100 cm, 100-150 cm, 150-200 cm (at late stage). In each sampling area, one sample quadrat was set. In each quadrant, the coverage and number of seedlings of Vallisneria natans, the height of mature plants, the length of the longest underground rhizomes, and the number of clones etc., were monitored respectively. Besides, the number of fruit and tuber produced by mature plants of Vallisneria natans was also monitored. Through the methods mentioned above, we could understand the response of typical functional traits of Vallisneria natans to different water depth conditions at different stages of the growth cycle (seed germination, growth and reproduction, etc.). Results show that seed germination and seedling survival rate were negatively correlated with water depth. In addition, with the increase of water depth, plant height, leaf length and specific leaf area increased significantly. Meanwhile, the aboveground biomass ratio of Vallisneria natans was the largest under the deepest water depth. Besides, sexual reproduction of Vallisneria natans was mainly carried out under shallow water condition (0-100 cm, >100-150 cm water depth), while clonal reproduction was mainly carried out by underground tuber under deep water condition (>150-200 cm water depth). The results highlight that the water depth had significant influence on the typical functional traits of Vallisneria natans in the stages of seed germination, growth and reproduction. This study can provide some theoretical support for the protection and recovery of Vallisneria natans population under water level fluctuation.
1 Germination indexes of seeds of Vallisneria natans under different water depth conditions不同水深条件下苦草种子的萌发指标 |
| 水深/cm | 盖度/% | 丛数/丛 | 叶片数/片 |
| 0~10 | 70 | 38 | 12 |
| >10~50 | 40 | 22 | 10 |
| >50~90 | 15 | 8 | 11 |
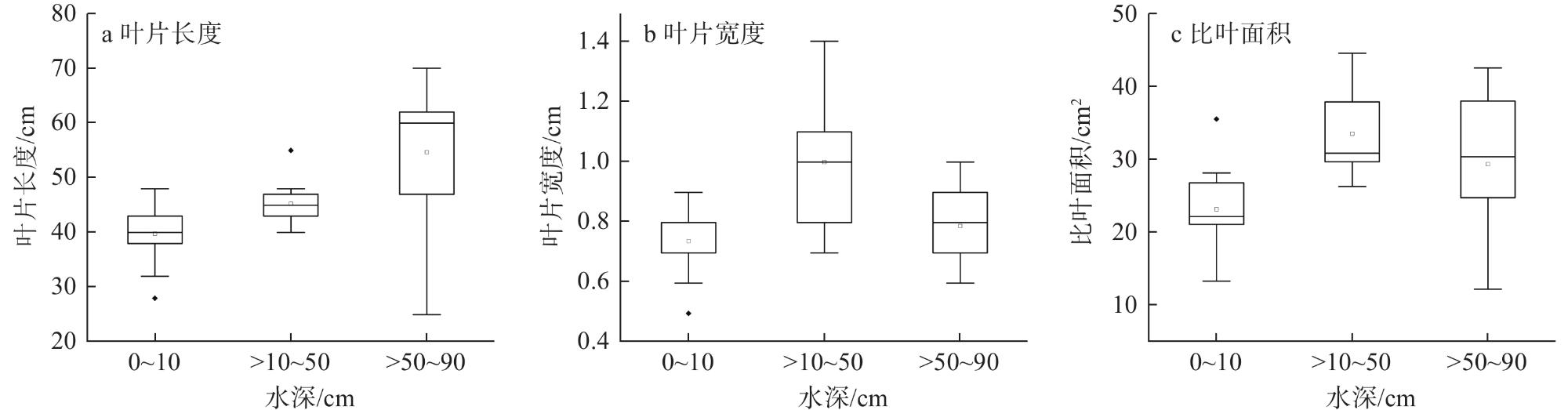
2 Influence of water depth on leaf length, leaf width, and specific leaf area of Vallisneria natans水深对苦草叶片长度、叶片宽度和比叶面积的影响 |
| 指标 | 单因素方差分析 | ||
| F | df | p | |
| 叶片长度 | 6.194 | 3 | 0.007 |
| 叶片宽度 | 5.361 | 3 | 0.012 |
| 比叶面积 | 3.931 | 3 | 0.033 |
1 Leaf length (a), leaf width (b) and specific leaf area (c) of Vallisneria natans under different water depth conditions不同水深条件下苦草的叶片长度(a)、叶片宽度(b)和比叶面积(c)
|
3 Proportion of aboveground biomass and belowground biomass of Vallisneria natans苦草地上生物量与地下生物量占比 |
| 水深/cm | 采样地1 | 采样地2 | 采样地3 | |||||
| 地下生物量占比/% | 地上生物量占比/% | 地下生物量占比/% | 地上生物量占比/% | 地下生物量占比/% | 地上生物量占比/% | |||
| 0~10 | 24.4 | 75.6 | 21.2 | 78.8 | 25.4 | 74.6 | ||
| >10~50 | 22.6 | 77.4 | 24.0 | 76.6 | 27.0 | 73.0 | ||
| >50~90 | 13.6 | 86.4 | 21.6 | 78.4 | 20.5 | 79.5 | ||
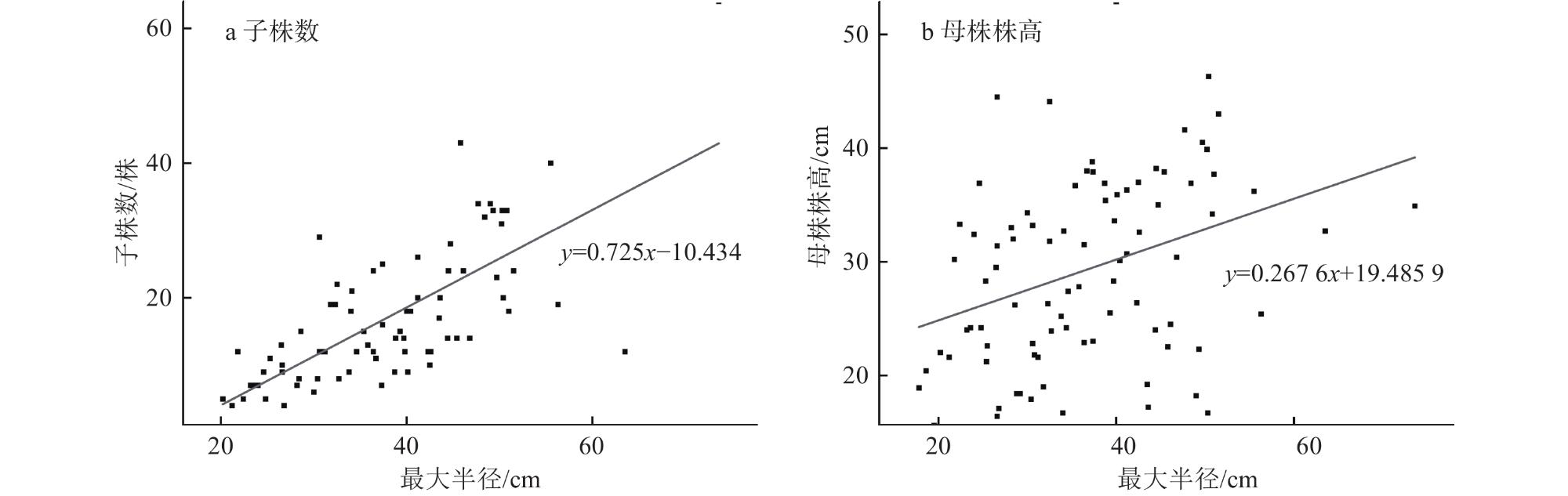
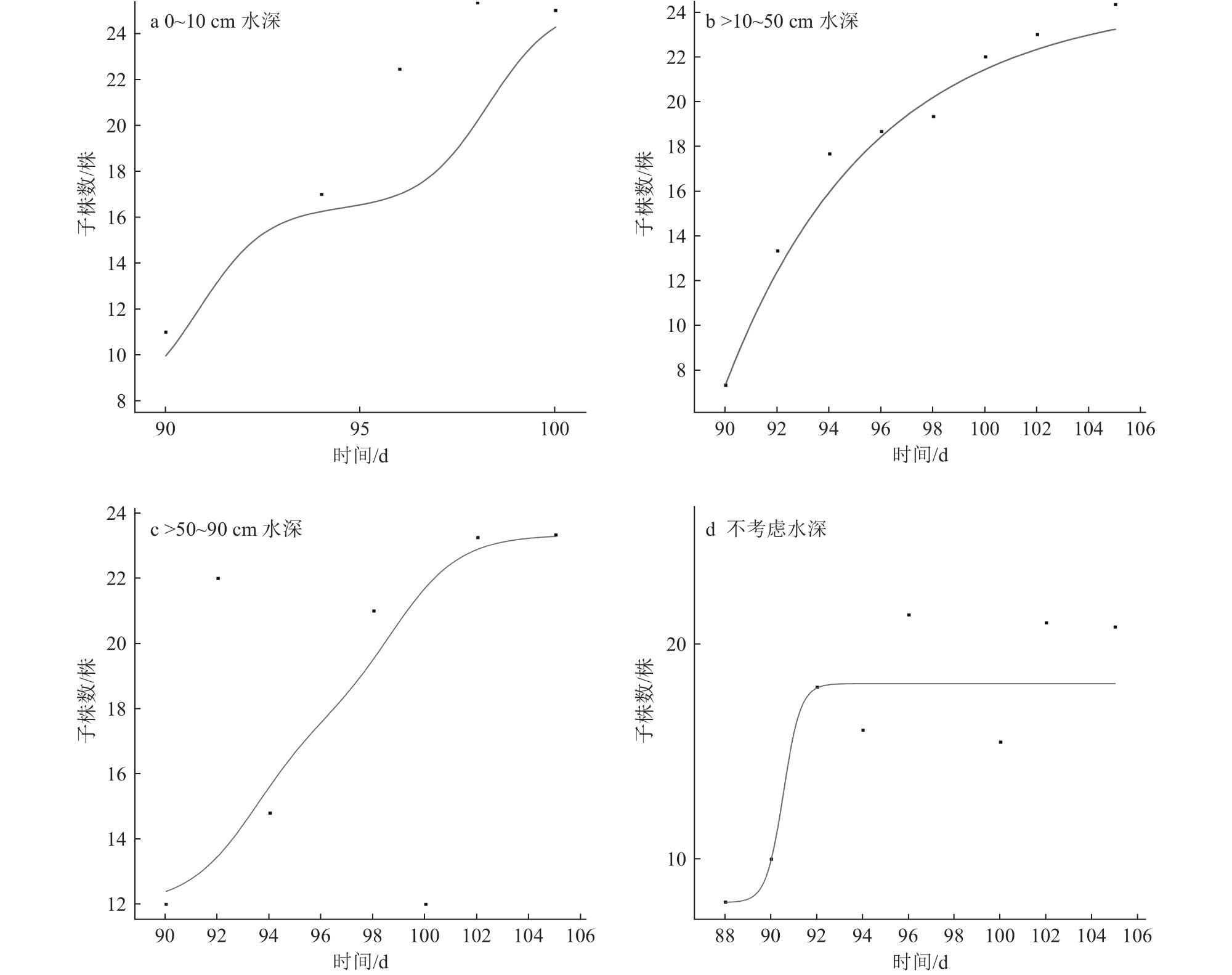
4 Influence of different water depth conditions on plant height, maximum radius and number of cloned plants of Vallisneria natans水深对苦草株高、最大半径和子株数的影响 |
| 单因素方差分析 | |||
| F值 | df值 | p值 | |
| 母株高 | 8.423 | 3 | <0.001 |
| 最大半径 | 8.451 | 3 | <0.001 |
| 子株数 | 3.184 | 3 | 0.03 |
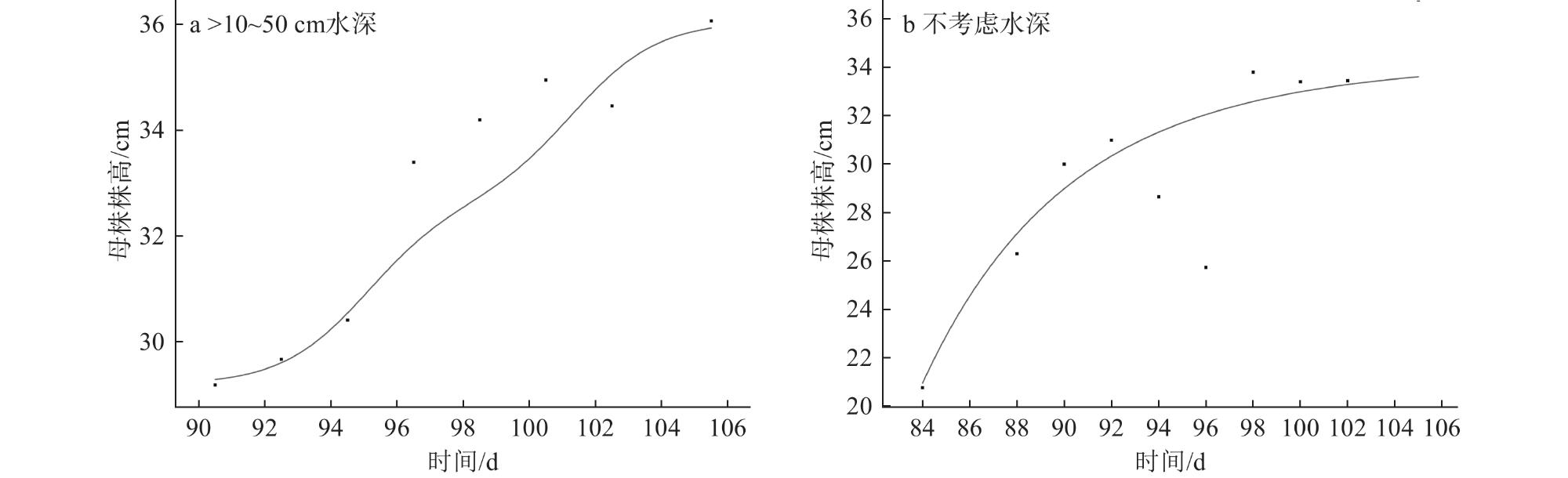
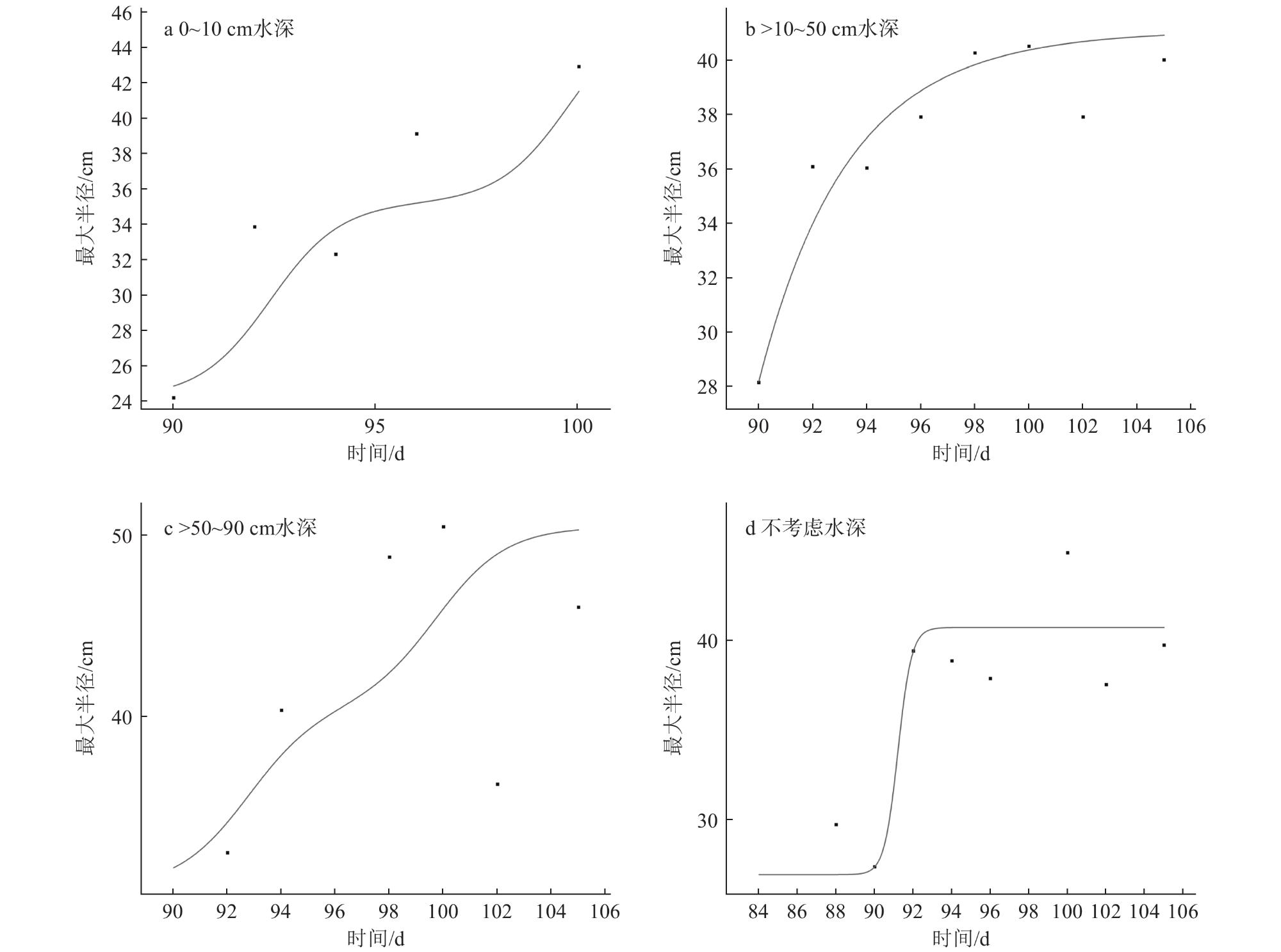
5 Plant height, maximum radius and number of cloned plants of Vallisneria natans under different water depth conditions不同水深条件下苦草的株高、最大半径和子株数 |
| 水深/cm | 母株高/cm | 最大半径/cm | 子株数/株 |
| 注:表中数据为平均值±标准误差。 | |||
| 0~10 | 28.55±2.11 | 36.04±0.91 | 16.13±1.27 |
| >10~50 | 31.37±2.02 | 38.46±0.96 | 17.31±1.36 |
| >50~90 | 35.95±2.11 | 40.88±0.96 | 19.34±1.27 |
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}