
草本植物群落的抗入侵性:影响机制与设计策略
|
李仓拴/男/博士/西北农林科技大学风景园林艺术学院讲师、硕士生导师/研究方向为城市生境与生物多样性营造、园林草本植物群落设计 |
|
张琼文/女/西北农林科技大学风景园林艺术学院在读硕士研究生/研究方向为植物群落设计 |
|
王菲/女/西北农林科技大学风景园林艺术学院在读硕士研究生/研究方向为植物群落设计 |
|
刘晖/女/博士/西安建筑科技大学建筑学院教授、博士生导师/本刊编委/研究方向为西北脆弱生态环境景观规划设计理论与方法、中国地景文化历史与理论 |
Copy editor: 李清清
收稿日期: 2023-12-31
修回日期: 2024-07-03
网络出版日期: 2025-12-16
基金资助
国家自然科学基金青年项目“园林草本植物群落CSR功能群构成与稳定关系研究”(编号32401654);陕西省自然科学基金青年项目“基于生物多样性的关中城市缀花草坪植物群落设计模式研究”(2021JQ-176)
版权
Invasion Resistance of Herbaceous Plant Community: Influence Mechanism and Design Strategy
|
LI Cangshuan, Ph.D., is a lecturer and master’s supervisor in the College of Landscape Architecture and Art, Northwest A&F University. His research focuses on ecological design for arid and semi-arid cities, and urban plant community design |
|
ZHANG Qiongwen is a master student in the College of Landscape Architecture and Art, Northwest A&F University. Her research focuses on plant community design |
|
WANG Fei is a master student in the College of Landscape Architecture and Art, Northwest A&F University. Her research focuses on plant community design |
|
LIU Hui, Ph.D., is a professor and doctoral supervisor in the College of Architecture, Xi’an University of Architecture and Technology, and an editorial board member of this journal. Her research focuses on theory and method of landscape planning and design for fragile ecological environments in northwest China, and history and theory of Chinese landscape culture |
Received date: 2023-12-31
Revised date: 2024-07-03
Online published: 2025-12-16
Copyright
除草一直是园林草本植物群落管理的棘手问题,而提高群落的抗入侵性是降低除草成本的关键途径。群落抗入侵机制作为入侵生态学研究的核心内容,旨在揭示生态因子对群落抗入侵性的影响规律。受制于研究范式的差异,这些规律须经过设计应用转换,才能真正实现理论指导实践。
通过文献分析,梳理群落抗入侵性影响机制,并从风景园林的视角提出群落抗入侵性提升策略。
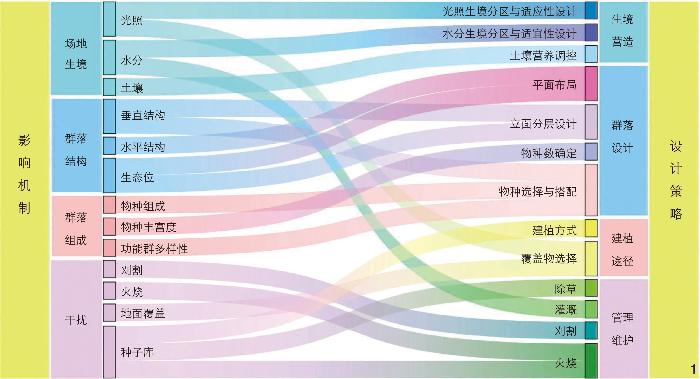
草本植物群落抗入侵性受生境、群落结构、群落组成和干扰等因素的综合影响,其中群落组成是主要影响要素;群落的抗入侵机制可通过生态学变量和设计变量的关联,转化为具体的生境营造、群落设计、建植途径和管理维护等提升抗入侵性的设计策略。
生态学基础理论可以被转换为对设计具有指导意义的操作方法,关键在于找到生态变量与设计变量的关联,这也是实现生态学与风景园林交叉的“桥梁”之一。
李仓拴 , 张琼文 , 王菲 , 刘晖 . 草本植物群落的抗入侵性:影响机制与设计策略[J]. 风景园林, 2024 , 31(11) : 79 -85 . DOI: 10.3724/j.fjyl.202309150424
Weed control has always been a difficult problem for urban green space managers around the world. Weeds grow rapidly, which not only compete with flowers for nutrients, water and other resources, but also serve as the intermediate host of a variety of pests and diseases. Therefore, the poor control of weeds will surely affect the growth of flowers and plants. The current weed control methods mainly include manual weed removal, mulch cultivation and use, chemical weed control, etc., some of which are time-consuming and laborious or prone to cause environmental pollution, thus failing to achieve efficient and ecological weed control. However, the biological control method is seldom used in garden green space, for which the main reason is that the survival mechanism between different species (such as adaptive relationship with the habitat, community structure, community composition, external interference, etc.) has not been transformed into operable rules and methods. The key to biological control is to improve the invasion resistance of plant community, which is also a key way to radically reduce the cost of weed control. Relevant research on the mechanism of invasion resistance, the core content of invasion ecology research, has revealed a large number of underlying rules. However, such research has neither systematically sorted out the influence mechanism on community invasion resistance nor designed the application and transformation of the aforesaid rules, making it difficult to really realize the value of guidance of invasion resistance for community design.
Through literature analysis, this research sorts out the influence mechanism of community invasion resistance, and puts forward relevant strategies for improving community invasion resistance from the perspective of landscape architecture. This research systematically analyzes the ecological factors affecting the invasion resistance of herbaceous plant community and their relationship with community invasion resistance in ecological research, and transforms them into the design strategies for improving community invasion resistance from the perspective of landscape architecture.
The invasion resistance of herbaceous plant community is influenced by a combination of factors such as habitat, community structure, community composition and disturbance, with community composition is the main influencing factor. Different factors have different impacts on the invasion resistance of herbaceous plant community, and communities composed of different species vary greatly in invasion resistance. 1) In terms of habitat, shady habitats perform best in invasion resistance, seconded by semi-aerobic habitats, while positive habitats perform worst; invasion is likely to occur in areas and years with relatively abundant rainfall; invasion is generally more likely to occur in fertile soils with relatively high carbon, nitrogen, and phosphorus contents. 2) In terms of community structure, increasing the number of community layers can help enhance the invasion resistance; when an invaded site has native species overlapping with the ecological niches of exotic species, it will be difficult for the exotic species to invade. 3) In terms of community composition, some special species in a plant community can increase the community’s invasion resistance; in general, the invasion resistance of a community is positively correlated with the number of species in the community; the higher the diversity of functional groups in a community, the stronger the community’s invasion resistance; theoretically, the edge area of a community is less resistant to disturbance, where the interface is prone to mutation, indicating that the edge area is less resistant to invasion. 4) In terms of disturbance, mowing can significantly reduce interspecific competition in a community, thus increasing the community’s invasion resistance; fire can reduce the seeds and propagules of weeds and inhibit the invasive disturbance of weed seeds on the community; gravel mulching can effectively reduce the germination of light-dependent weed seeds in the soil; and the fewer the species in the seed bank and the seed rain, the lower the possibility of community invasion.
Therefore, the mechanism of community invasion resistance can be transformed into specific design strategies for invasion resistance enhancement such as habitat creation, community design, pathway establishment, and management and maintenance through the association of ecological and design variables. By creating herbaceous plant community from the four aspects of site habitat, community structure, community composition and disturbance relationship, the invasion resistance of a community can be effectively improved, so that the community can inhibit or influence the growth of surrounding weeds through its own “immune” system, without introducing chemicals that are difficult to degrade into the system or causing environmental problems related to chemical pesticides, while reducing the production cost, which can help promote the sustainable development of urban herbaceous plant community. Further, this research confirms that the basic theories of ecology can be converted into design-guided methods, and the key lies in the association between ecological variables and design variables, which is one of the “bridges” to realize the intersection of ecology and landscape architecture. However, this research decomposes the complex community system into single factors and simply considers the single-factor effect on species, which makes the research limited to a certain extent. In the future, it is supposed to take into account more composite factor variables to elucidate the mechanism of species coexistence and community invasion resistance, so as to better solve the problem of weeds in urban gardens.
| [1] |
郑景明, 马克平. 植物群落多样性与可入侵性关系研究进展[J]. 应用生态学报, 2006, 17 (7): 1338-1343
ZHENG J M, MA K P. Research Advances in the Relationships Between Biodiversity and Invasiveness Within Plant Communities[J]. Chinese Journal of Applied Ecology, 2006, 17 (7): 1338-1343
|
| [2] |
万方浩, 侯有明, 蒋明星. 入侵生物学[M]. 北京: 科学出版社, 2015.
WAN F H, HOU Y M, JIANG M X. Invasion Biology[M]. Beijing: Science Press, 2015.
|
| [3] |
黄建华, 周晓然. 《北京市城镇绿地养护管理投资标准的意见》实施北京城镇绿地养护费提高[J]. 绿化与生活, 2022 (3): 28-29
HUANG J H, ZHOU X R. Implementation of the Opinions on Investment Standards for Urban Green Space Maintenance and Management in Beijing Has Led to an Increase in Maintenance Fees for Urban Green Spaces in Beijing[J]. Greening and Life, 2022 (3): 28-29.
|
| [4] |
国务院办公厅印发《关于科学绿化的指导意见》[J].新疆林业, 2021(6): 14-17.
Guidance on the Greening of Science Issued by China Academic Journal Electronic Publishing House[J]. Xinjiang Forestry, 2021(6): 14-17.
|
| [5] |
CHATTO B, STEVEN W. Beth Chatto’s Gravel Garden[M]. Suffolk: Antique Collectors Club Ltd, 2000.
|
| [6] |
SCHMITHALS A, KÜHN N. To Mulch or not to Mulch? Effects of Gravel Mulch Toppings on Plant Establishment and Development in Ornamental Prairie Plantings[J]. PloS One, 2017, 12 (2): 35-67.
|
| [7] |
BARR S, JONAS J L, PASCHKE M W. Optimizing Seed Mixture Diversity and Seeding Rates for Grassland Restoration[J]. Restoration Ecology, 2017, 25 (3): 396-404
|
| [8] |
JASINSKAITE S, PILIPAVICIUS V, LAZAUSKAS P. Perennial Weed Control by Two Layer Ploughing[J]. Agronomy Research, 2009 (7): 277-282.
|
| [9] |
GIORIA M, O’FLYNN C, OSBORNE B A. A Review of the Impacts of Major Terrestrial Invasive Alien Plants in Ireland[J]. Biology & Environment Proceedings of the Royal Irish Academy, 2018, 118 (3): 157-179.
|
| [10] |
刘晖, 王晶懋, 吴小辉. 生境营造的实验性研究[J]. 中国园林, 2017, 33 (3): 19-23
LIU H, WANG J M, WU X H. An Experimental Research Approach on the Habitat-Site Design[J]. Chinese Landscape Architecture, 2017, 33 (3): 19-23
|
| [11] |
徐汝梅, 叶万辉. 生物入侵: 理论与实践[M]. 北京: 科学出版社, 2003.
XU R M, YE W H. Biological Invasion: Theory and Practice[M]. Beijing: Science Press, 2003.
|
| [12] |
象伟宁, 王涛, 汪辉. 魅力的巴斯德范式 vs.盛行的玻尔范式: 谁是生态系统服务研究中更具生态实践智慧的研究范式?[J]. 现代城市研究, 2018 (7): 2-6,19
XIANG W N, WANG T, WANG H. Pasteur’s Quadrant: An Appealing Ecophronetic Alternative to the Prevalent Bohr’s Quadrant in Ecosystem Services Research[J]. Landscape Ecology, 2018 (7): 2-6,19.
|
| [13] |
万方浩, 谢丙炎, 杨国庆. 入侵生物学[M]. 北京: 科学出版社, 2011.
WAN F H, XIE B Y, YANG G Q. Invasion Biology[M]. Beijing: Science Press, 2011.
|
| [14] |
李仓拴. 城市建成环境地被植物群落设计模式研究: 西安建筑科技大学实验与实践[D]. 西安: 西安建筑科技大学, 2019.
LI C S. Research on the Design Model of Urban Built up Environment Ground Cover Plant Community-Experimental and Practical Study at Xi’an University of Architecture and Technology[D]. Xi’an: Xi’an University of Architecture and Technology, 2019.
|
| [15] |
DAVIS M A, GRIME P, THOMPSON K. Fluctuating Resources in Plant Communities: A General Theory of Invasibility[J]. Journal of Ecology, 2000, 88 (3): 528-534
|
| [16] |
STOHLGREN T J, BULL K A, OTSUKI Y, et al. Riparian Zones as Havens for Exotic Plant Species in the Central Grasslands[J]. Plant Ecology, 1998, 138 (13): 113-125.
|
| [17] |
LARSON D L, ANDERSON P J, NEWTON W. Alien Plant Invasion in Mixed-Grass Prairie: Effects of Vegetation Type and Anthropogenic Disturbance[J]. Ecological Applications, 2001, 11 (1): 128-141
|
| [18] |
HOBBS R J, YATES S, MOONEY H A. Long-Term Data Reveal Complex Dynamics in Grassland in Relation to Climate and Disturbance[J]. Ecological Monographs, 2007, 77 (4): 545-568
|
| [19] |
WEDIN D A, TILMAN D. Influence of Nitrogen Loading and Species Composition on the Carbon Balance of Grasslands[J]. 1996, 274(5293): 1720-1723.
|
| [20] |
RENNE I J, TRACY B F, COLONNA I A. Shifts in Grassland Invasibility: Effects of Soil Resources, Disturbance, Composition, and Invader Size[J]. Ecology, 2006, 87 (9): 2264-2277
|
| [21] |
HITCHMOUGH J. Sowing Beauty: Designing Flowering Meadows from Seed[M]. Portland: Timber Press, 2017.
|
| [22] |
WEST C. Planting in a Post-Wild World: Designing Plant Communities for Resilient Landscapes[M]. Portland: Timber Press, 2015.
|
| [23] |
NAEEM S, KNOPS J M H, TILMAN D, et al. Plant Diversity Increases Resistance to Invasion in the Absence of Covarying Extrinsic Factors[J]. Oikos, 2000, 91 (1): 97-108.
|
| [24] |
周宇峰, 周国模. 斑块边缘效应的研究综述[J]. 华东森林经理, 2007, 21 (2): 1-8
ZHOU Y F, ZHOU G M. Summary of Research on Edge Effect of Patch[J]. East China Forest Management, 2007, 21 (2): 1-8
|
| [25] |
DARWIN C. The Origin of Species by Means of Natural Selection[M]. London: John Murray, 1859.
|
| [26] |
ELTON C S. The Ecology of Invasions by Animals and Plants[M]. London: Butler and Tanner Ltd., 1958.
|
| [27] |
FARGIONE J, BROWN C S, TILMAN D. Community Assembly and Invasion: An Experimental Test of Neutral Versus Niche Processes[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100 (15): 8916-8920.
|
| [28] |
RAINER T, WEST C. Planting in a Post-Wild World: Designing Plant Communities for Resilient Landscapes[M]. Timber Press, 2015.
|
| [29] |
LI S P, JIA P, FAN S Y, et al. Functional Traits Explain the Consistent Resistance of Biodiversity to Plant Invasion Under Nitrogen Enrichment[J]. Ecology Letters, 2021, 25 (4): 778-789.
|
| [30] |
郑景明, 马克平. 入侵生态学[M]. 北京: 高等教育出版社, 2010.
ZHENG J M, MA K P. Invasion Ecology[M]. Beijing: Higher Education Press, 2010.
|
| [31] |
KENNEDY T A, NAEEM S, HOWE K M, et al. Biodiversity as a Barrier to Ecological Invasion[J]. Nature, 2002, 417: 636-638
|
| [32] |
WISERS K, ALLEN R B, CLINTON P W, et al. Community Structure and Forest Invasion by an Exotic Herb over 23 Years[J]. Ecology, 1998, 79: 2071-2081
|
| [33] |
姚向阳, 李委涛, 郑玉龙. 群落组成及物种间相互作用对外来植物入侵的影响[J]. 生态学杂志, 2014, 33 (7): 1953-1959
YAO X Y, LI W T, ZHENG Y L. Effect of Community Composition and Species Interactions on the Invasion of Alien Plants[J]. Chinese Journal of Ecology, 2014, 33 (7): 1953-1959.
|
| [34] |
WANG C Y, YU Y L, CHENG H Y, et al. Which Factor Contributes Most to the Invasion Resistance of Native Plant Communities Under the Co-invasion of Two Invasive Plant Species?[J]. Science of the Total Environment, 2022, 813: 152628.
|
| [35] |
马文红, 方精云. 中国北方典型草地物种丰富度与生产力的关系[J]. 生物多样性, 2006 (1): 21-28
MA W H, FANG J Y. The Relationship Between Species Richness and Productivity in Four Typical Grasslands of Northern China[J]. Biodiversity Science, 2006 (1): 21-28
|
| [36] |
孙国钧, 张荣, 周立. 植物功能多样性与功能群研究进展[J]. 生态学报, 2003 (7): 1430-1435
SUN G J, ZHANG R, ZHOU L. Trends and Advances in Researches on Plant Functional Diversity and Functional Groups[J]. Acta Ecologica Sinica, 2003 (7): 1430-1435
|
| [37] |
BYUN C, DE BLOIS S, BRISSON J. Plant Functional Group Identity and Diversity Determine Biotic Resistance to Invasion by an Exotic Grass[J]. Journal of Ecology, 2013, 101 (1): 128-139
|
| [38] |
BYUN C, DE BLOIS S, BRISSON J. Restoring Functionally Diverse Communities Enhances Invasion Resistance in a Freshwater Wetland[J]. Journal of Ecology, 2020, 108 (6): 2485-2498
|
| [39] |
MASON T J, KRISTINE F, JOLLEY D F, et al. Functional Richness and Identity Do not Strongly Affect Invasibility of Constructed Dune Communities[J]. Plos One, 2017, 12 (1): 351-367.
|
| [40] |
PRIEUR-RICHARD A H, LAVOREL S, GRIGULIS K, et al. Plant Community Diversity and Invasibility by Exotics: Invasion of Mediterranean Old Fields by Conyza Bonariensis and Conyza Canadensis[J]. Ecology Letters, 2010, 3 (5): 412-422.
|
| [41] |
国家牧草产业技术体系. 中国栽培草地[M]. 北京: 科学出版社, 2015.
National Grassland Industry Technology System. Cultivated grasslands in China[M]. Beijing: Science Press, 2015.
|
| [42] |
姚忠, 吴永明, 游海林, 等. 竞争-耐胁迫-杂草型植物对策理论及其应用研究进展[J]. 生态学报, 2022, 42 (1): 24-36
YAO Z, WU Y M, YOU H L, et al. Research Progress of Plant Competitor-Stress Tolerator-Ruderal (CSR) Theory and Its Application[J]. Acta Ecologica Sinica, 2022, 42 (1): 24-36
|
| [43] |
YANG Z P, MINGGAGUD H, WANG Q, et al. Interacting Effects of Nitrogen Addition and Mowing on Plant Diversity and Biomass of a Typical Grassland in Inner Mongolia[J]. Agronomy, 2023, 13 (8): 2125.
|
| [44] |
吉普森.禾草和草地生态学 [M].北京: 高等教育出版社, 2018.
GIBSON D J. Grass and Grassland Ecology[M]. Beijing: Higher Education Press, 2018.
|
| [45] |
LECK M A, PARKER V T, SIMPSON R L. Ecology of Soil Seed Bank[M]. Cambridge, MA: Academic Press, 1989: 385-444.
|
| [46] |
于顺利, 蒋高明. 土壤种子库的研究进展及若干研究热点[J]. 植物生态学报, 2003 (4): 552-560
YU S L, JIANG G M. The Research Development of Soil Seed Bank and Several Hot Topics[J]. Acta Phytoecologica Sinica, 2003 (4): 552-560
|
| [47] |
李仓拴, 刘晖, 杨伊婷, 等. 西北干旱城市破碎化绿地生境的植物群落设计途径研究[J]. 风景园林, 2019, 26 (2): 88-93
LI C S, LIU H, YANG Y T, et al. Plant Community Design Approaches for Fragmented Green Space Habitats in Northwest China Arid Cities[J]. Landscape Architecture, 2019, 26 (2): 88-93.
|
| [48] |
王晶懋, 刘晖, 吴小辉, 等. 基于场地小气候特征的草本植物群落设计研究[J]. 风景园林, 2018, 25 (4): 98-102
WANG J M,LIU H,WU X H,et al. A Research on Herbaceous Plant Community Fabric Design Based on Site Microclimate Characteristic[J]. Landscape Architecture, 2018, 25 (4): 98-102.
|
| [49] |
刘晖, 李莉华, 徐鼎黄, 等. 自然环境条件影响下的西北城市绿地生境营造途径[J]. 西安建筑科技大学学报(自然科学版), 2016, 48 (4): 556-561
LIU H, LI L H, XU D H, et al. Approach for Urban Green Space Site-Habitat Design and Construction Method Based on the Natural Environment Conditions in North-West China[J]. Journal of Xi’an University of Architecture & Technology (Natural Science Edition), 2016, 48 (4): 556-561.
|
| [50] |
HITCHMOUGH J, WAGNER M, AHMAD H, et al. Extended Flowering and High Weed Resistance Within Two Layer Designed Perennial “Prairie-Meadow” Vegetation[J]. Urban Forestry & Urban Greening, 2017, 27: 117-126.
|
| [51] |
冉海燕, 兰献敏, 陈仕红, 等.果园以草治草研究进展[J].农技服务, 2019, 36(2): 68, 70.
RAN H Y, LAN X M, CHEN S H, et al. Research Progress on Grass Control in Orchards[J]. Agricultural Technical Services, 2019, 36(2): 68, 70.
|
| [52] |
许凯扬, 叶万辉. 群落可入侵性及环境胁迫[J]. 热带亚热带植物学报, 2003, 11 (1): 75-82
XU K Y, YE W H. Community Invasibility and Environmental Stress[J]. Journal of Tropical and Subtropical Botany, 2003, 11 (1): 75-82
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}