
|
王安琪(1999-),女,硕士研究生,主要从事野生动植物保护与利用研究. E-mail: hanguang2021@126.com |
收稿日期: 2024-03-01
修回日期: 2024-06-22
网络出版日期: 2026-03-11
基金资助
新疆自然保护地调查与国家公园潜力区科学考察(2021xjkk1201)
Vegetation community diversity and spatial distribution pattern in Ganjiahu Saxoul National Nature Reserve, Xinjiang
Received date: 2024-03-01
Revised date: 2024-06-22
Online published: 2026-03-11
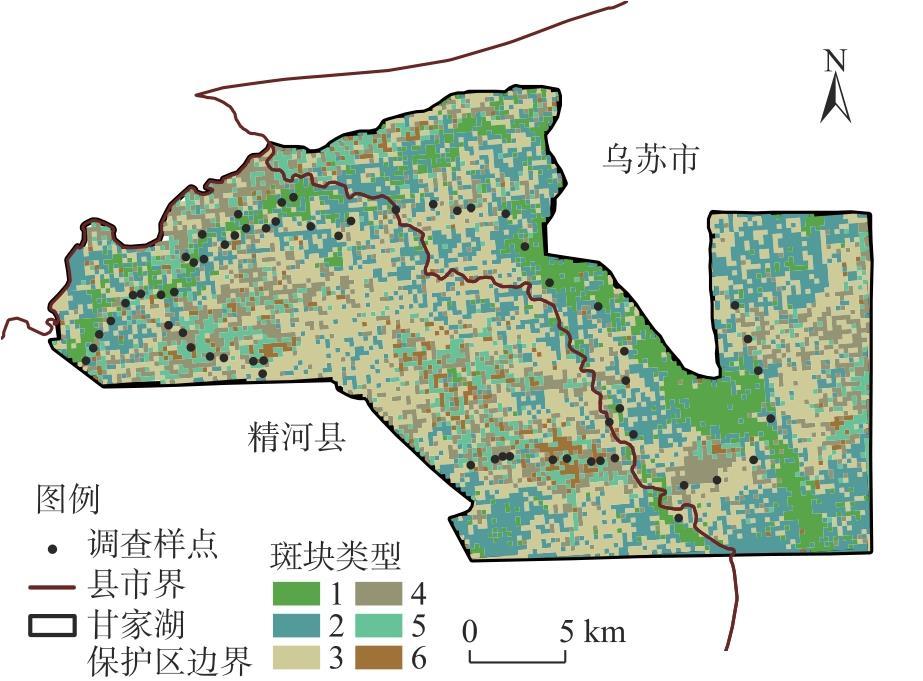
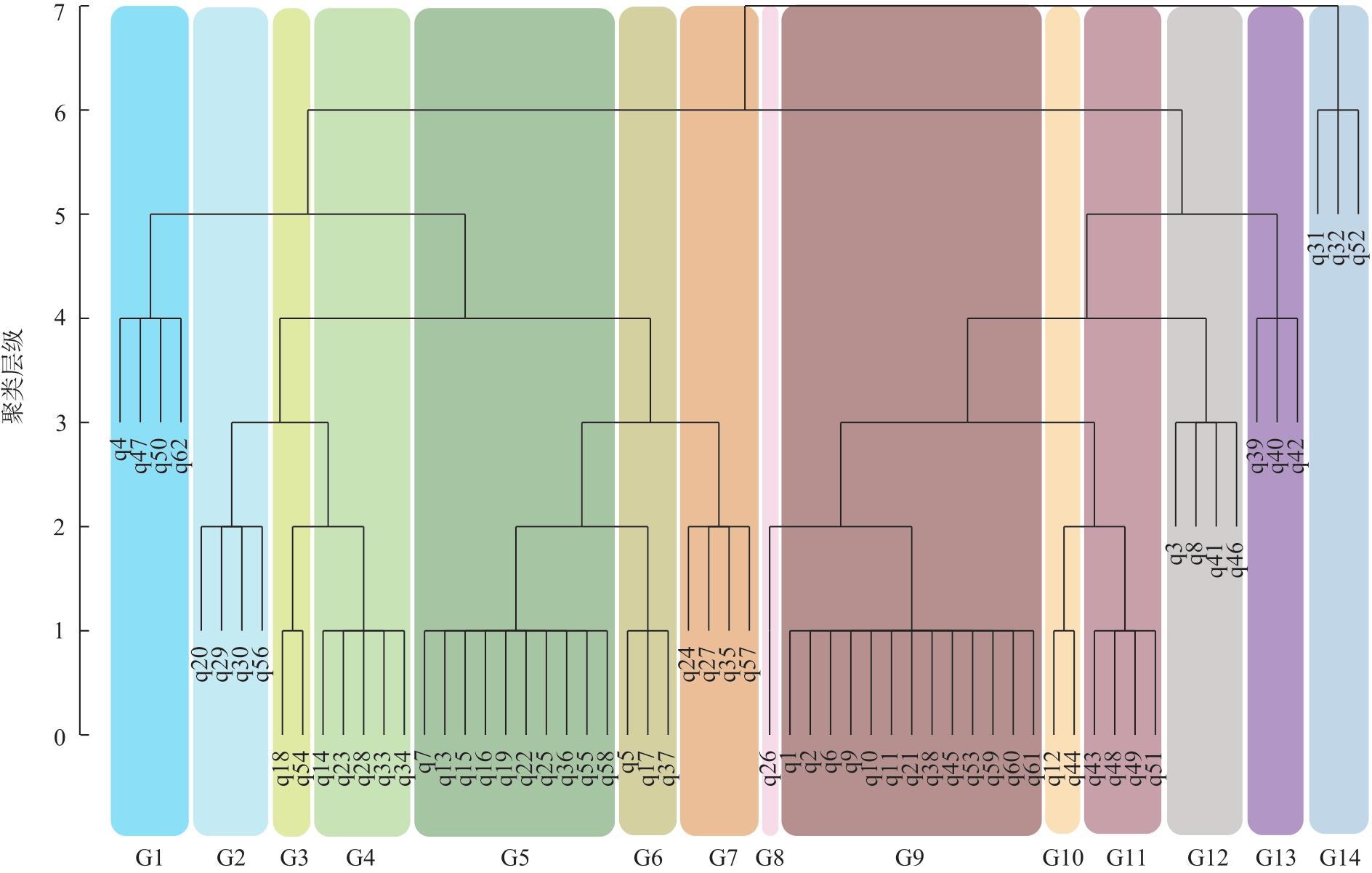
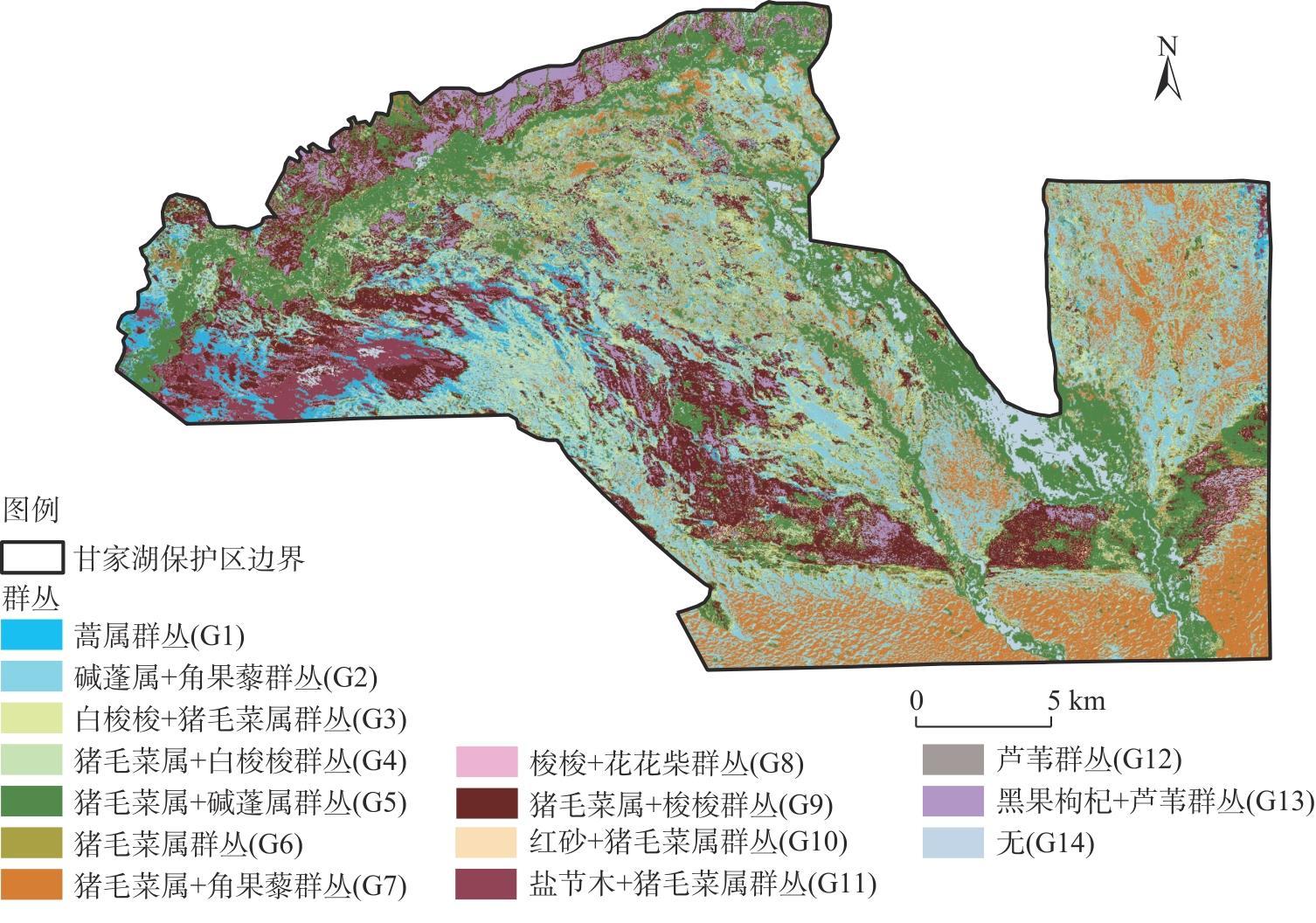
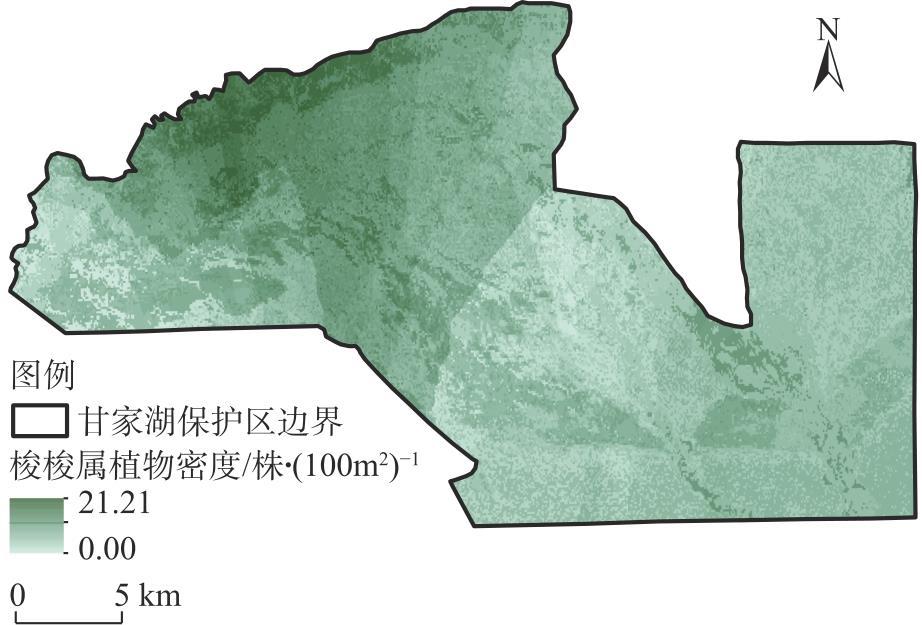
新疆甘家湖梭梭林国家级自然保护区位于典型的绿洲-荒漠过渡带,明确保护区内主要植被类型及分布有助于生物多样性的保护与恢复。以2022年同期遥感影像为样点布设依据,在2023年8月进行实地调查,利用双向指示种分析法(TWINSPAN)对保护区内植被样方进行分类,计算各植被类型的α多样性指数,随后构建随机森林模型以评估植被样方的聚类结果并可视化。结果表明:(1)在甘家湖保护区,草本植物以猪毛菜属(Salsola spp.)分布最为广泛,灌木以梭梭(Haloxylon ammodendron)、碱蓬属(Suaeda spp.)、红砂(Reaumuria songarica)、角果藜(Ceratocarpus arenarius)等为这一地区主要的植被组成。(2)TWINSPAN将62个植被样方划分成14个类型,其中猪毛菜属+碱蓬属群丛(Ass. Salsola spp.+Suaeda spp.,G5)所占面积最大,且植被多样性最高;猪毛菜属+角果藜群丛(Ass. Salsola spp.+Ceratocarpus arenarius,G7)物种多样性较高,同时面积占比较大;单一植被型群丛(G14)Shannon-Wiener多样性指数与Simpson优势度指数均为最低,Pielou均匀度指数为1,在保护区中主要体现为农田和盐渍化极高的裸地。(3)随机森林模型的总体精度达到87.10%,Kappa系数达到0.8553,错分误差与漏分误差相对较小。(4)梭梭和白梭梭集中分布在保护区西北部核心区以及中偏南部的缓冲区。研究结果可为保护区后续精准管理奠定基础。
王安琪 , 张钰 , 刘佳伟 , 赵威 , 刘馨雨 , 李凯 . 新疆甘家湖梭梭林国家级自然保护区植被群落多样性及空间分布格局[J]. 干旱区地理, 2024 , 47(11) : 1841 -1851 . DOI: 10.12118/j.issn.1000-6060.2024.133
The Ganjiahu Saxoul National Nature Reserve in Xinjiang, China, is situated in a typical oasis-desert transition zone. Clearly identifying the main vegetation types and their distribution within the reserve is essential for biodiversity protection and restoration. Based on remote sensing images from 2022, a field survey was conducted in August 2023. Vegetation samples from the Ganjiahu Saxoul National Nature Reserve were classified using the two-way indicator species analysis (TWINSPAN) method. Alpha diversity indices of each vegetation type were calculated, followed by the development of a random forest model to evaluate and visualize the clustering results of vegetation quadrats. The results are as follows: (1) In the Ganjiahu Saxoul National Nature Reserve, the dominant herbaceous plants are primarily Salsola spp., while the principal shrubs include Haloxylon ammodendron, Suaeda spp., Reaumuria songarica, and Ceratocarpus arenarius. (2) Sixty-two vegetation sample plots were categorized into 14 types in TWINSPAN. Among these, the Ass. Salsola spp.+Suaeda spp. (G5) covered the largest area and exhibited the highest vegetation diversity. The Ass. Salsola spp. + Ceratocarpus arenarius (G7) showed both high species diversity and extensive coverage. The G14 type represented a single vegetation community with the lowest Shannon-Wiener and Simpson’s diversity indices and had a Pielou’s evenness index of 1, predominantly comprising farmland and highly saline bare land in the Ganjiahu Saxoul National Nature Reserve. (3) The random forest model achieved an overall accuracy of 87.10%, with a Kappa coefficient of 0.8553, and demonstrated relatively low commission and omission errors. (4) H. ammodendron and H. persicum are mainly distributed in the core area in the northwest and the buffer zone extending from the central to southern parts of the Ganjiahu Saxoul National Nature Reserve. These findings provide a foundation for future precise management of the Ganjiahu Saxoul National Nature Reserve.
表1 甘家湖保护区主要植被概况Tab. 1 Overview of the main vegetation in Ganjiahu Nature Reserve |
| 序号 | 植物种类 | 拉丁学名 | 平均高度/cm | 平均盖度/% | 平均重要值 |
|---|---|---|---|---|---|
| 1 | 猪毛菜属 | Salsola spp. | 12.49 | 9.80 | 31.02 |
| 2 | 梭梭 | Haloxylon ammodendron | 195.74 | 42.39 | 9.70 |
| 3 | 碱蓬属 | Suaeda spp. | 30.24 | 17.26 | 8.27 |
| 4 | 芦苇 | Phragmites australis | 45.59 | 8.29 | 6.68 |
| 5 | 红砂 | Reaumuria songarica | 49.53 | 19.66 | 4.88 |
| 6 | 角果藜 | Ceratocarpus arenarius | 6.66 | 7.80 | 4.67 |
| 7 | 多枝柽柳 | Tamarix ramosissima | 166.40 | 54.40 | 4.44 |
| 8 | 蒿属 | Artemisia spp. | 42.52 | 23.87 | 4.40 |
| 9 | 盐节木 | Halocnemum strobilaceum | 19.85 | 27.46 | 4.33 |
| 10 | 戈壁藜 | Iljinia regelii | 31.83 | 6.78 | 3.30 |
| 11 | 黑果枸杞 | Lycium ruthenicum | 23.13 | 19.88 | 3.23 |
| 12 | 白梭梭 | Haloxylon persicum | 131.08 | 25.96 | 2.99 |
| 13 | 骆驼刺 | Alhagi camelorum | 41.89 | 14.33 | 2.45 |
| 14 | 花花柴 | Karelinia caspia | 39.00 | 5.55 | 1.26 |
| 15 | 小果白刺 | Nitraria sibirica | 30.38 | 26.50 | 1.11 |
| 16 | 胡杨 | Populus euphratica | 235.00 | 45.00 | 0.93 |
| 17 | 驼绒藜 | Krascheninnikovia ceratoides | 53.11 | 13.89 | 0.75 |
| 18 | 铃铛刺 | Caragana halodendron | 133.00 | 33.40 | 0.73 |
| 19 | 沙拐枣属 | Calligonum spp. | 37.60 | 17.40 | 0.72 |
| 20 | 盐爪爪 | Kalidium foliatum | 29.60 | 9.00 | 0.66 |
| 21 | 盐穗木 | Halostachys caspica | 78.75 | 47.00 | 0.45 |
| 22 | 雾冰藜 | Grubovia dasyphylla | 3.00 | 1.67 | 0.44 |
| 23 | 抱茎独行菜 | Lepidium perfoliatum | 41.00 | 9.17 | 0.39 |
| 24 | 大叶补血草 | Limonium gmelinii | 51.83 | 9.67 | 0.38 |
| 25 | 芨芨草 | Neotrinia splendens | 78.75 | 18.25 | 0.36 |
| 26 | 甘草 | Glycyrrhiza uralensis | 25.00 | 20.00 | 0.28 |
| 27 | 白麻 | Apocynum pictum | 90.00 | 17.00 | 0.26 |
| 28 | 东方旱麦草 | Eremopyrum orientale | 8.00 | 10.00 | 0.24 |
| 29 | 沙蓬 | Agriophyllum pungens | 29.50 | 3.00 | 0.20 |
| 30 | 蓝刺头 | Echinops sphaerocephalus | 16.00 | 1.50 | 0.18 |
| 31 | 肉苁蓉 | Cistanche deserticola | 18.00 | 1.00 | 0.13 |
| 32 | 盐生假木贼 | Anabasis salsa | 13.50 | 4.00 | 0.13 |
| 33 | 荠 | Capsella bursa-pastoris | 6.00 | 1.00 | 0.04 |
表2 不同植被群丛α多样性指数Tab. 2 α diversity indices of different vegetation clusters |
| 群丛编号 | 样方数量 | Shannon-Wiener 多样性指数 | Simpson优势度 | Pielou均匀度指数 |
|---|---|---|---|---|
| G1 | 4 | 1.354 | 0.594 | 0.565 |
| G2 | 4 | 1.643 | 0.745 | 0.685 |
| G3 | 2 | 1.475 | 0.652 | 0.640 |
| G4 | 5 | 1.661 | 0.774 | 0.668 |
| G5 | 10 | 1.864 | 0.798 | 0.645 |
| G6 | 3 | 1.437 | 0.727 | 0.739 |
| G7 | 4 | 1.748 | 0.792 | 0.703 |
| G8 | 1 | 1.304 | 0.700 | 0.810 |
| G9 | 13 | 1.339 | 0.664 | 0.558 |
| G10 | 2 | 1.409 | 0.716 | 0.677 |
| G11 | 4 | 1.593 | 0.760 | 0.641 |
| G12 | 4 | 1.782 | 0.775 | 0.658 |
| G13 | 3 | 1.305 | 0.706 | 0.729 |
| G14 | 3 | 0.693 | 0.500 | 1.000 |
图3 基于随机森林模型的甘家湖保护区植被类型分布Fig. 3 Distribution of vegetation types in Ganjiahu Nature Reserve based on random forest model |
表3 基于随机森林模型划分的各植被类型面积及精度验证Tab. 3 Verification of the area and accuracy of each vegetation type based on random forest model |
| 群丛编号 | 面积/km2 | 面积占比/% | 生产者精度/% | 使用者精度/% |
|---|---|---|---|---|
| G1 | 28.50 | 4.64 | 75.00 | 100.00 |
| G2 | 116.15 | 18.91 | 100.00 | 80.00 |
| G3 | 18.45 | 3.00 | 100.00 | 100.00 |
| G4 | 59.31 | 9.66 | 60.00 | 75.00 |
| G5 | 124.87 | 20.33 | 90.00 | 69.23 |
| G6 | 26.07 | 4.25 | 100.00 | 100.00 |
| G7 | 83.71 | 13.63 | 100.00 | 100.00 |
| G8 | 2.84 | 0.46 | 100.00 | 100.00 |
| G9 | 81.26 | 13.23 | 84.62 | 91.67 |
| G10 | 6.63 | 1.08 | 100.00 | 66.67 |
| G11 | 17.05 | 2.78 | 100.00 | 100.00 |
| G12 | 1.72 | 0.28 | 75.00 | 100.00 |
| G13 | 32.28 | 5.26 | 100.00 | 100.00 |
| G14 | 15.39 | 2.51 | 66.67 | 100.00 |
感谢新疆甘家湖梭梭林国家级自然保护区管理局精河分局、乌苏分局对于本研究的大力支持,感谢保护区五道泉管护站工作人员的付出与帮助。
| [1] |
方精云, 郭柯, 王国宏, 等. 《中国植被志》的植被分类系统、植被类型划分及编排体系[J]. 植物生态学报, 2020, 44(2): 96-110.
[
|
| [2] |
|
| [3] |
刘炳强. 甘家湖荒漠林区环境恶化原因及治理对策[J]. 防护林科技, 2014(10): 85, 126.
[
|
| [4] |
陈雨婷, 马松梅, 张丹, 等. 新疆同域分布梭梭和白梭梭多样性格局及其形成机制[J]. 植物生态学报, 2024, 48(1): 56-67.
[
|
| [5] |
俞阗, 张君萍, 马洪雨, 等. 梭梭抗逆相关研究进展[J]. 新疆农业科学, 2009, 46(4): 907-912.
[
|
| [6] |
蔡新斌, 吴俊侠. 甘家湖自然保护区白梭梭种群特征与动态分析[J]. 干旱区资源与环境, 2016, 30(7): 90-94.
[
|
| [7] |
李引湣. 近三十年来新疆甘家湖自然保护区梭梭林退化过程研究[D]. 乌鲁木齐: 新疆师范大学, 2013.
[
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
文铜, 柳小妮, 纪童, 等. 基于植被指数的三江源高寒草地植物分类与识别方法研究[J]. 草地学报, 2022, 30(7): 1811-1818.
[
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
王国宏, 方精云, 郭柯, 等. 《中国植被志》研编内容与规范[J]. 植物生态学报, 2020, 44(2): 128-178.
[
|
| [23] |
刘丽燕, 蔡新斌, 江晓珩, 等. 甘家湖梭梭林国家级自然保护区植被类型和特征[J]. 防护林科技, 2016(6): 54-57.
[
|
| [24] |
马倩, 楚新正, 孙慧兰, 等. 新疆甘家湖梭梭林绿洲-荒漠过渡带土地利用/覆被动态变化分析[J]. 新疆师范大学学报(自然科学版), 2014, 33(1): 8-13, 37.
[
|
| [25] |
王育松, 上官铁梁. 关于重要值计算方法的若干问题[J]. 山西大学学报(自然科学版), 2010, 33(2): 312-316.
[
|
| [26] |
|
| [27] |
李俊晓, 李朝奎, 殷智慧. 基于ArcGIS的克里金插值方法及其应用[J]. 测绘通报, 2013(9): 87-90.
[
|
| [28] |
刘丽燕, 蔡新斌, 江晓珩, 等. 新疆甘家湖梭梭林自然保护区植物组成及地理成分分析[J]. 植物科学学报, 2016, 34(5): 695-704.
[
|
| [29] |
高晓宇, 郝海超, 张雪琪, 等. 中国西北干旱区植被水分利用效率变化对气象要素的响应——以新疆为例[J]. 干旱区地理, 2023, 46(7): 1111-1120.
[
|
| [30] |
|
| [31] |
徐先英, 严平, 郭树江, 等. 干旱荒漠区绿洲边缘典型固沙灌木的降水截留特征[J]. 中国沙漠, 2013, 33(1): 141-145.
[
|
| [32] |
|
| [33] |
李引湣, 曲宁. 甘家湖梭梭林的变化浅析[J]. 和田师范专科学校学报, 2014(5): 77-82.
[
|
| [34] |
朱家龙, 周智彬, 王立生, 等. 免灌人工梭梭林生长与土壤水分变化的耦合关系[J]. 干旱区地理, 2022, 45(5): 1579-1590.
[
|
| [35] |
曹家睿. 甘家湖白梭梭空间格局分析[J]. 南方农业, 2020, 14(27): 186-188, 218.
[
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}