
Received date: 2024-06-12
Revised date: 2024-08-14
Online published: 2026-03-12
Copyright
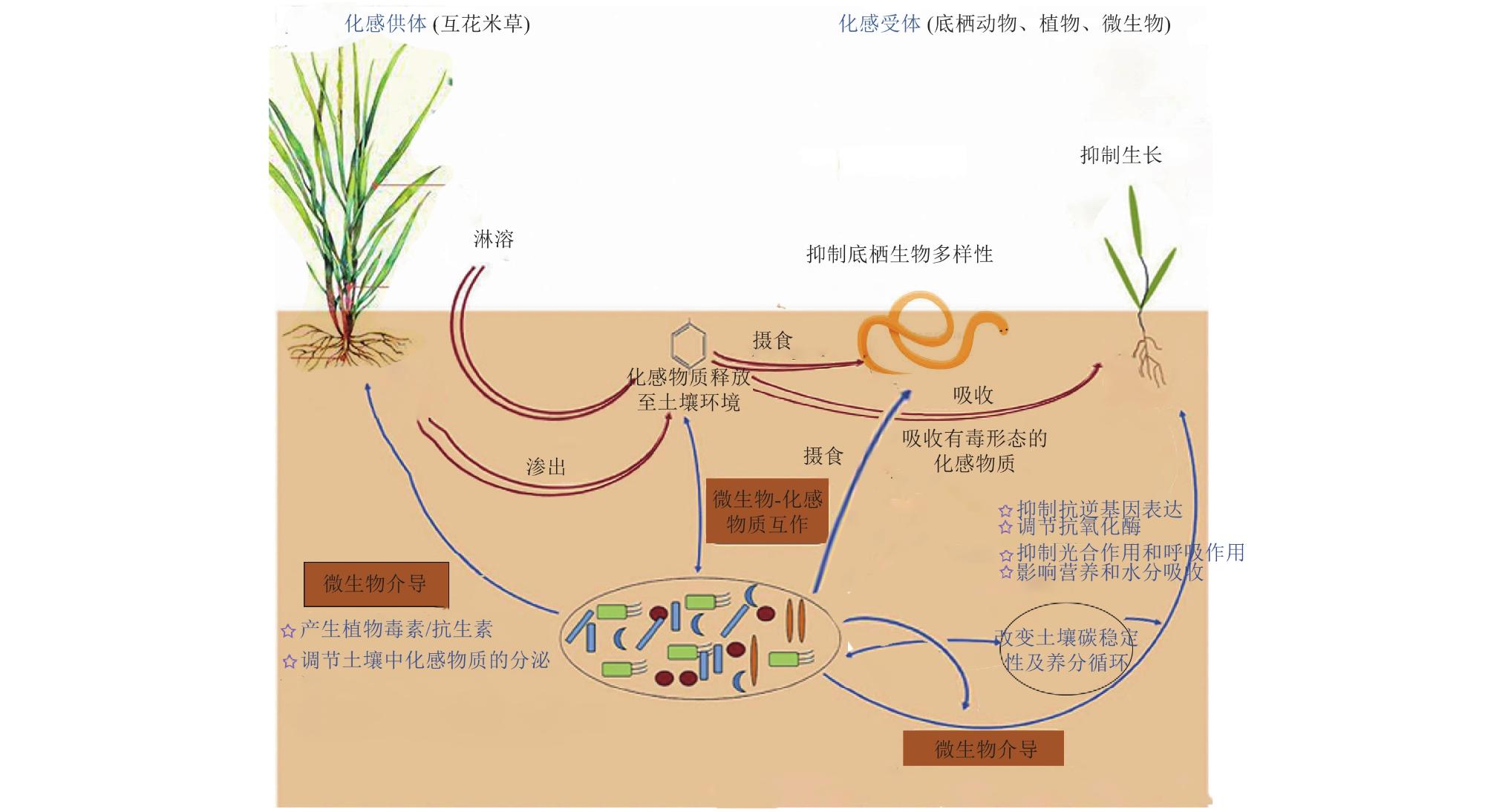
Spartina alterniflora is a widely recognized invasive species that poses significant threats to native ecosystems. Its successful invasion is closely linked to its allelopathic effects, which involve the release of chemical compounds that influence the growth and physiological processes of neighboring organisms. This review comprehensively summarizes the classification of allelochemicals produced by Spartina alterniflora and explores the multiple mechanisms through which its allelopathic effects operate. The allelopathic substances secreted by Spartina alterniflora include organic acids, flavonoids, coumarins, alkaloids, and other secondary metabolites, primarily released through root exudates, leaching, and decomposition of plant residues. These compounds interfere with nutrient and water uptake in native plants by disrupting root function, thus limiting their access to essential growth resources. Moreover, Spartina alterniflora allelochemicals inhibit photosynthesis and respiration by damaging chloroplast ultrastructure and reducing photosynthetic pigments, stomatal conductance, and transpiration rates in native plants such as mangroves and herbaceous species. Additionally, these chemicals affect enzyme activities and cell membrane permeability, altering antioxidant enzyme systems (e.g., superoxide dismutase and peroxidase) and increasing lipid peroxidation markers, thereby compromising plant physiological function. The allelopathic effects extend beyond plants, impacting soil microbial communities by reducing the colonization of beneficial mycorrhizal fungi and altering microbial biomass and nutrient cycling processes, particularly carbon and nitrogen dynamics. These microbial changes negatively affect soil health and nutrient availability, further influencing plant community structure.The invasion of Spartina alterniflora also disrupts benthic animal communities in coastal wetlands, leading to declines in species diversity and richness of mollusks and nematodes. This is partly attributed to chemical changes in the sediment environment induced by allelochemicals released into the rhizosphere and litterfall, which alter food web dynamics and nutrient cycling in these habitats.Interestingly, Spartina alterniflora exhibits autotoxicity, where some allelochemicals produced inhibit its own seed germination and seedling growth, affecting its population dynamics. However, most research has focused on its heterospecific allelopathic interactions, and further studies are required to elucidate the mechanisms and implications of its autotoxicity.The allelopathic interaction between Spartina alterniflora and native plants is bidirectional. Certain native species such as Phragmites australis (common reed) and mangroves release compounds that suppress Spartina alterniflora growth, indicating complex chemical interactions that could be leveraged for ecological management. In addition to ecological impacts, Spartina alterniflora allelochemicals have practical applications. Extracts from this species demonstrate inhibitory effects on harmful algal blooms, such as Microcystis aeruginosa and Prorocentrumdonghaiense, through oxidative stress induction and disruption of photosynthetic systems. Moreover, flavonoids isolated from Spartina alterniflora possess anti-inflammatory properties and have been explored for cosmetic applications, suggesting potential economic value.Future research should broaden the scope to include effects on lower plants, fungi, soil biochemical properties, and animals, and investigate environmental factors influencing allelochemical production and release. Advanced analytical methods are needed to isolate and identify allelochemicals, while genetic and molecular studies could clarify regulatory mechanisms. Exploring integrated management strategies combining biological, chemical, and physical methods is essential to control Spartina alterniflora invasions while harnessing its beneficial properties. Overall, this review provides a comprehensive understanding of the allelopathic roles of Spartina alterniflora in invaded ecosystems, offering critical insights for ecological restoration and invasive species management.
Wen Jiao , Chen Yujun , Li Mei , Deng Chuangfa , Yin Xiaoyang , Liang Weishen , Li Jun . Progress in allelopathy research of Spartina alterniflora[J]. Wetland Science, 2025 , 23(6) : 1300 -1309 . DOI: 10.13248/j.cnki.wetlandsci.20240161
1 Summary of the allelopathic effects of Spartina alterniflora on plants, animals, and microorganisms互花米草化感作用对植物、动物和微生物的影响 |
| 1 |
| 2 |
| 3 |
| 4 |
| 5 |
| 6 |
| 7 |
| 8 |
| 9 |
| 10 |
| 11 |
| 12 |
| 13 |
| 14 |
| 15 |
| 16 |
| 17 |
| 18 |
| 19 |
| 20 |
| 21 |
| 22 |
| 23 |
| 24 |
| 25 |
| 26 |
| 27 |
| 28 |
| 29 |
| 30 |
| 31 |
| 32 |
| 33 |
| 34 |
| 35 |
| 36 |
| 37 |
| 38 |
| 39 |
| 40 |
| 41 |
| 42 |
| 43 |
| 44 |
| 45 |
| 46 |
| 47 |
| 48 |
| 49 |
| 50 |
| 51 |
| 52 |
| 53 |
| 54 |
| 55 |
| 56 |
| 57 |
| 58 |
| 59 |
| 60 |
| 61 |
| 62 |
| 63 |
| 64 |
| 65 |
| 66 |
| 67 |
| 68 |
| 69 |
| 70 |
| 71 |
| 72 |
| 73 |
| 74 |
| 75 |
| 76 |
| 77 |
| 78 |
| 79 |
| 80 |
| 81 |
| 82 |
| 83 |
| 84 |
| 85 |
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}