
Impact of environmental interfaces on the decomposition characteristics of typical hydrophyte litters in Baiyangdian Lake
Received date: 2024-05-06
Revised date: 2024-07-02
Online published: 2026-03-12
Copyright
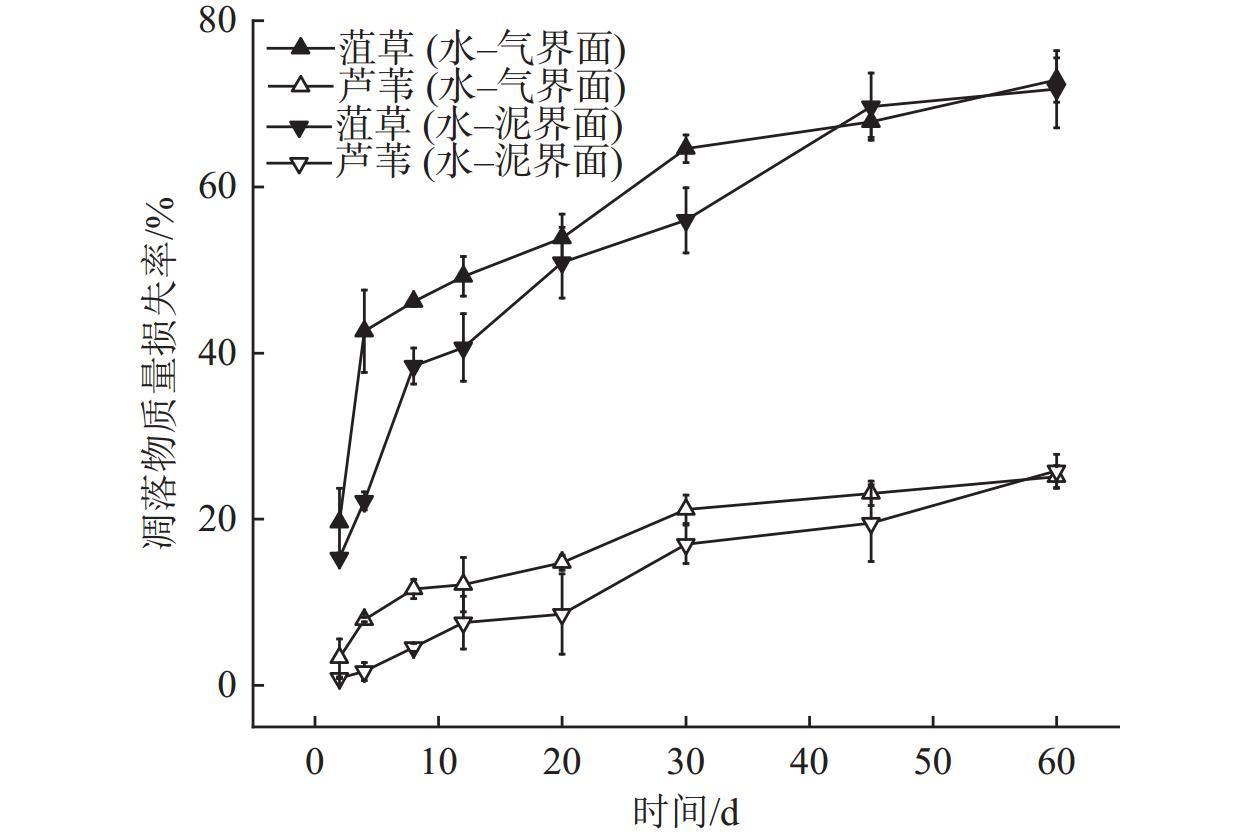
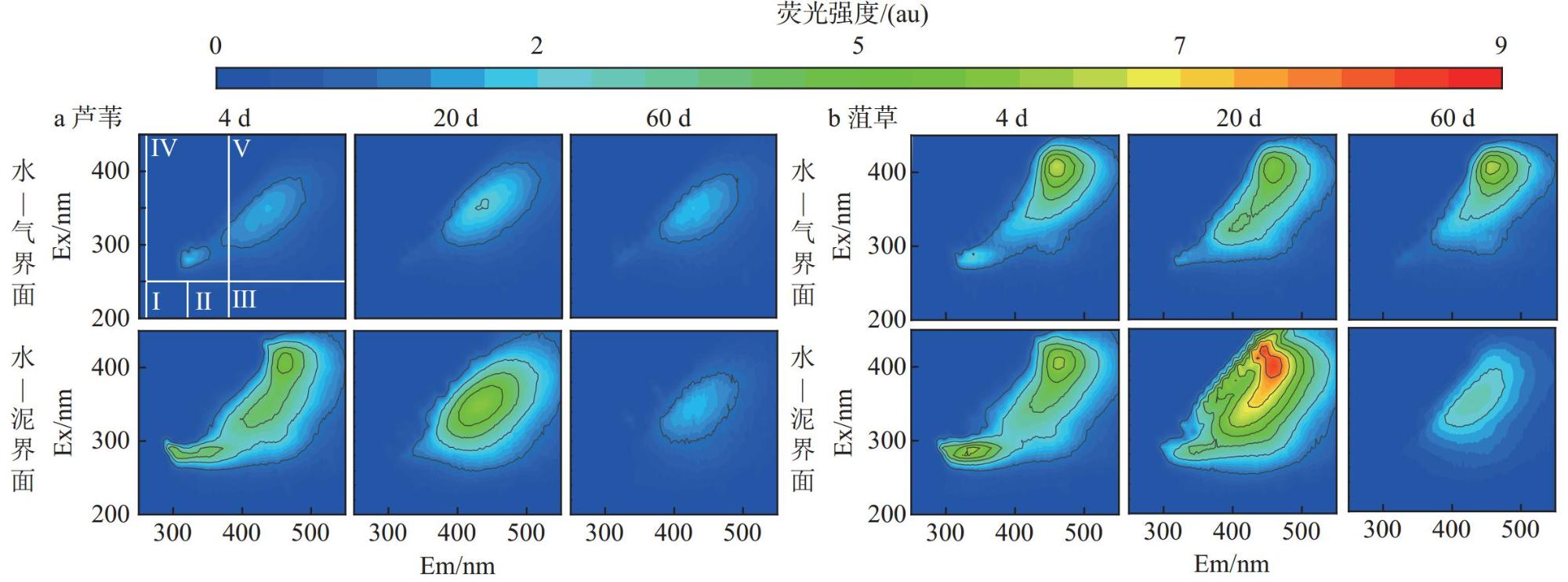
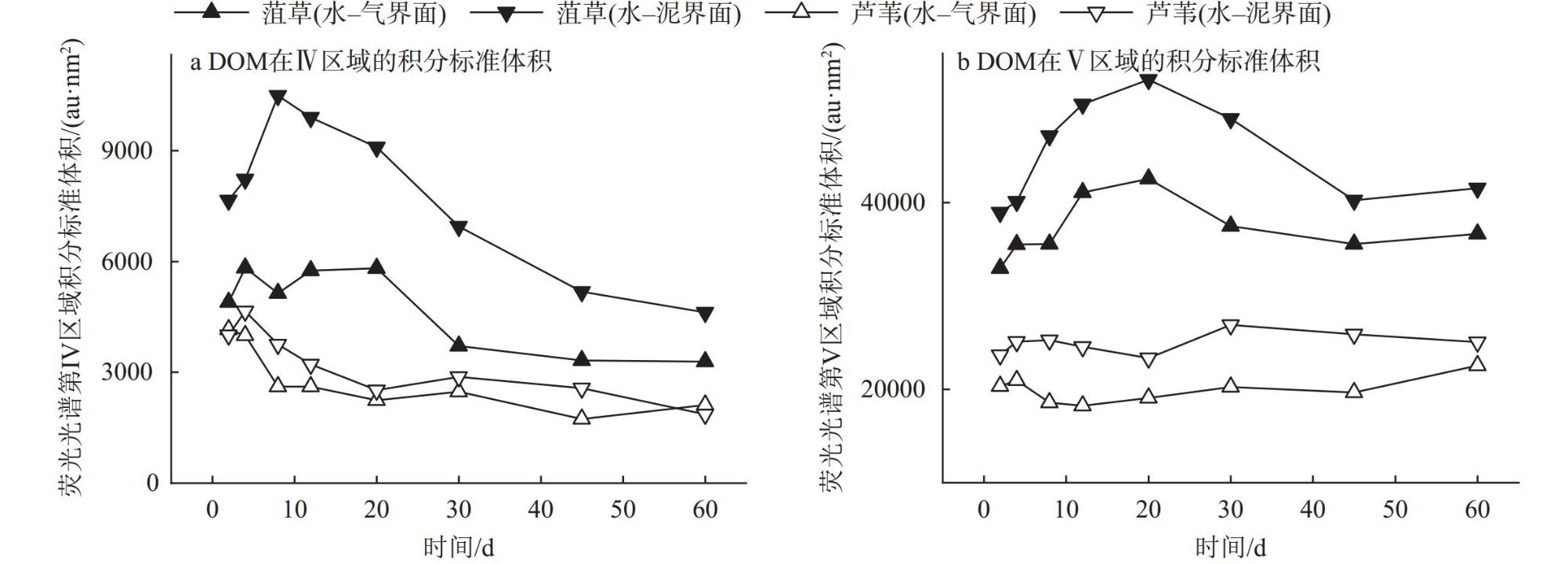
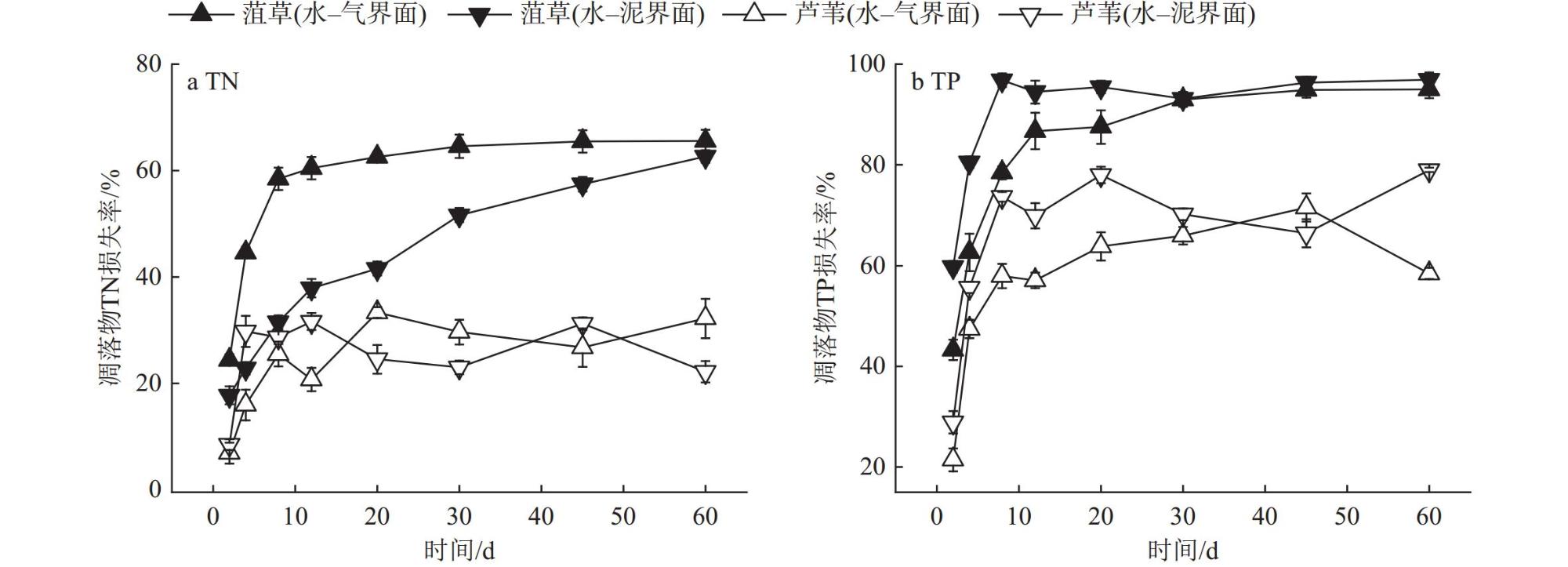
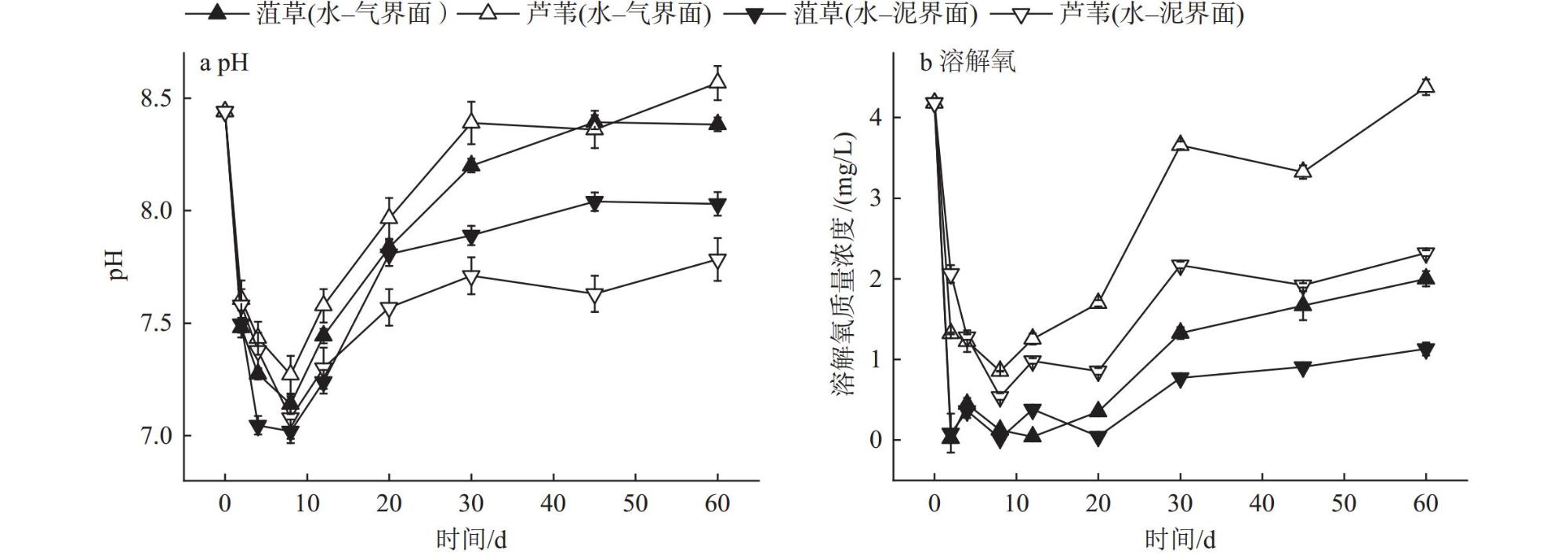
Interfaces are critical locations where abrupt changes in wetland environmental conditions occur, exerting significant impacts on wetland carbon-sink. In order to investigate the impact of environmental interfaces on the decomposition of plant litter, we selected Potamogeton crispus and Phragmites australis, the typical hydrophyte in Baiyangdian Lake, as research objects. Using litter bag method to investigate the decomposition dynamics of these two plant litters under the treatment conditions of the water-gas interface and water-sediment interface. Additionally, we also analyzed the release characteristics of dissolved organic matter (DOM), nitrogen, and phosphorus from two tyres of plant litter during the decomposition process, as well as the changes in pH value and dissolved oxygen (DO) content in the decomposition water bodies.The results showed that within the first 30 days of decomposition, the mass loss rates of Potamogeton crispus and Phragmites australis litters at the water-gas interface were higher than those at the water-sediment interface, thereafter, the mass loss rates of the two plant litters at the two interfaces gradually became similar. At both interfaces, the mass loss rate and decomposition rate of Potamogeton crispus litter were higher than those of Phragmites australis litter. The results of regional integration and fluorescence spectral indices indicate that the DOM released from the decomposition of Potamogeton crispus and Phragmites australis litters was mainly composed of dissolved microbial metabolites and humic-like substances, with distinct autochthonous characteristics. The changes in DOM fluorescence signal intensity at the water-gas interface were more gradual than those at the water-sediment interface for both plant litters. According to the results of parallel factor analysis (PARAFAC), Potamogeton crispus litter released four types of DOM components (tryptophan-like (C1), terrestrial humic-like (C2 and C3), fulvic-like (C4)) at the water-sediment interface, and three components (C1, C2, and C4) were identified at the water-gas interface. For Phragmites australis litter, only two components (C2 and C4) were separated at both interfaces. The release of nitrogen and phosphorus from the two litters was mainly concentrated in the early decomposition stage (0-12 days), and the N and P loss rates of Potamogeton crispus litter were higher than those of Phragmites australis litter, while the release patterns of nitrogen and phosphorus from each type of plant litter were similar at the two interfaces. The pH value and DO content in the every decomposition water body decreased rapidly in the 0-8 days, and then gradually recovered, with the recovery rate at the water-gas interface being faster than that at the water-sediment interface.
Wang Zhiwei , Zhang Kegang , Ben Xianglong , Yan Zhaoyi , Yuan Chungang . Impact of environmental interfaces on the decomposition characteristics of typical hydrophyte litters in Baiyangdian Lake[J]. Wetland Science, 2025 , 23(6) : 1288 -1299 . DOI: 10.13248/j.cnki.wetlandsci.20240122
1 The Olson index model for the mass loss of various plant litters at the water-gas interface and water-sediment interface各植物凋落物在水–气界面和水–泥界面质量损失的Olson指数模型 |
| 植物 | 界面 | Olson拟合方程 | k | R2 |
| 注: R2为Olson拟合方程方差。 | ||||
| 菹草 | 水–气 | Mt =0.7167e−0.018t | 0.018 | 0.755 6 |
| 水–泥 | Mt =0.8229e−0.020t | 0.020 | 0.905 9 | |
| 芦苇 | 水–气 | Mt =0.9462e−0.005t | 0.005 | 0.881 2 |
| 水–泥 | Mt =0.9956e−0.005t | 0.005 | 0.981 0 | |
2 Fluorescence index (FI), biological index (BIX), and humification index (HIX) of DOM released from plant litters decomposition植物凋落物分解释放DOM的荧光指数(FI)、生物指数(BIX)和腐殖化指数(HIX) |
| 界面 | 时间/d | 菹草 | 芦苇 | |||||
| FI | HIX | BIX | FI | HIX | BIX | |||
| 水–气 | 4 | 2.29 | 0.50 | 1.04 | 1.98 | 0.57 | 1.00 | |
| 20 | 2.11 | 0.71 | 0.94 | 1.89 | 0.61 | 0.87 | ||
| 60 | 2.30 | 0.77 | 0.91 | 2.01 | 0.76 | 0.90 | ||
| 水–泥 | 4 | 2.08 | 0.52 | 0.92 | 2.10 | 0.53 | 0.93 | |
| 20 | 2.37 | 0.68 | 0.89 | 2.03 | 0.83 | 0.88 | ||
| 60 | 2.14 | 0.88 | 0.88 | 1.91 | 0.90 | 0.83 | ||
| 1 |
| 2 |
| 3 |
| 4 |
| 5 |
| 6 |
| 7 |
| 8 |
| 9 |
| 10 |
| 11 |
| 12 |
| 13 |
| 14 |
| 15 |
| 16 |
| 17 |
| 18 |
| 19 |
| 20 |
| 21 |
| 22 |
| 23 |
| 24 |
| 25 |
| 26 |
| 27 |
| 28 |
| 29 |
| 30 |
| 31 |
| 32 |
| 33 |
| 34 |
| 35 |
| 36 |
| 37 |
| 38 |
| 39 |
| 40 |
| 41 |
| 42 |
| 43 |
| 44 |
| 45 |
| 46 |
| 47 |
| 48 |
| 49 |
| 50 |
| 51 |
| 52 |
| 53 |
| 54 |
| 55 |
| 56 |
| 57 |
| 58 |
| 59 |
| 60 |
| 61 |
| 62 |
| 63 |
| 64 |
| 65 |
| 66 |
| 67 |
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}