
Characteristics and influencing factors of plant community changes along elevational gradients in the marshland of Shengjin Lake
Received date: 2024-01-24
Revised date: 2024-04-05
Online published: 2026-03-12
Copyright
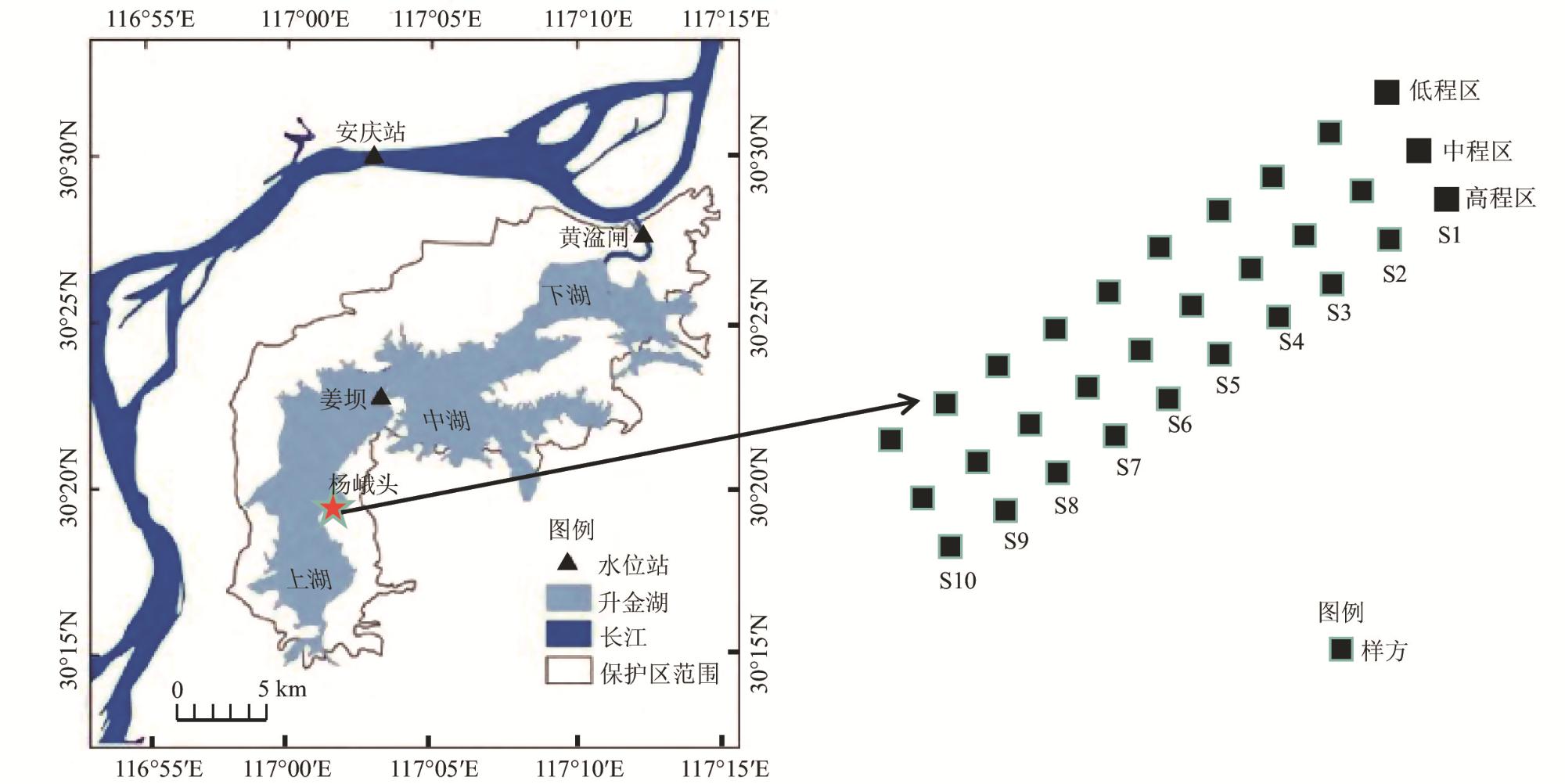
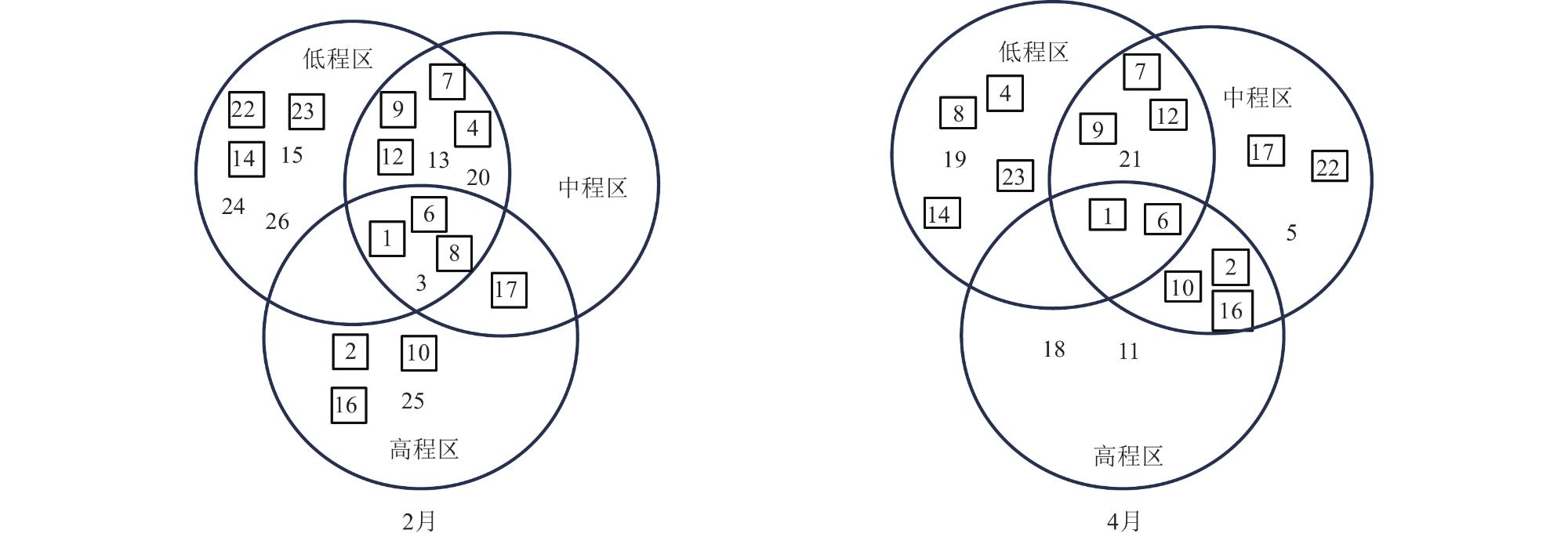
To reveal the distribution patterns of wetland plant communities across elevational gradients, a combined approach utilizing sampling lines and plots was employed to investigate the plant communities along these gradients in the Shengjin Lake marshland at different seasons. The results showed that the flora of the Shengjin Lake marshland comprised a total of 26 herbaceous plant species, belonging to 15 families and 24 genera, Asteraceae and Poaceae were the dominant families, with Carex thunbergia as the dominant species. Annual and biennial plants accounted for a relatively high proportion of 69.2%. The community structure in the high and middle elevations exhibited similarities, with a notable difference from the low elevation area. Carex thunbergii was the dominant species in all three elevations during both seasons. However, the main associated species differed significantly among the elevations, and there were also some differences across different seasons. Plant species gradually changed from species adapted to long-term flooding to species adapted to short-term flooding, there were few common species among the three areas, with only 4 common species in February and 2 in April. From February to April, the increased flooding in the low elevation area posed greater challenges for plant survival. Additionally, the heterogeneity of the intertidal environment intensified, resulting in the number of common species in the low and middle elevation areas, from 10 to 6. For species diversity, as the elevation gradient increased, the total species count in each area exhibited a general decreasing trend. Specifically, in February, there was a significant difference (p<0.01) in the average number of species between the low elevation area and the other two areas. Due to environmental heterogeneity, the other diversity indices did not consistently reflect a trend corresponding to changes in species richness. From February to April, the average number of species and Shannon index in the low elevation area sample plots decreased significantly (p<0.05), whereas the changes in the high elevation area were not significant. Based on the results, the factors influencing the characteristics of plant communities were analyzed and discussed, with water level fluctuations identified as the primary factor affecting the distribution heterogeneity and diversity of plant communities in the Shengjin Lake marshland.
Ou Zulan , Yang Xingxing , Li Jiaxin , Xue Yuanyuan , Chen Wenwen , Wang Guangyao , Li Chunlin , Wang Lingling , Chen Yansong . Characteristics and influencing factors of plant community changes along elevational gradients in the marshland of Shengjin Lake[J]. Wetland Science, 2025 , 23(3) : 505 -515 . DOI: 10.13248/j.cnki.wetlandsci.20240035
1 Species composition, important values and life forms of plants in the Shengjin Lake marshland升金湖洲滩植物物种组成及重要值与生活型 |
| 科/属/种 | 生活型 | 重要值 | ||||||
| 2月 | 4月 | |||||||
| 高程区 | 中程区 | 低程区 | 高程区 | 中程区 | 低程区 | |||
| 注:a、b、p分别表示一年生草本(annual herb)、二年生草本(biennial herb)、多年生草本(perennial herb)植物。表中无重要值的物种在该区域未出现。 | ||||||||
| 一 莎草科Cyperaceae | ||||||||
| (一)薹草属Carex | ||||||||
| 1陌上菅Carex thunbergii | p | 35.08 | 32.15 | 28.91 | 49.22 | 60.53 | 45.4 | |
| 二 禾本科Poaceae | ||||||||
| (二)芦苇属Phragmites | ||||||||
| 2芦苇Phragmites australis | p | 30.8 | 19.39 | 6.29 | ||||
| (三)虉草属Phalaris | ||||||||
| 3虉草Phalaris arundinacea | p | 5.17 | 10.26 | 3.35 | ||||
| (四)早熟禾属Poa | ||||||||
| 4早熟禾Poa annua | a | 8.64 | 4.9 | 4.34 | ||||
| (五)披碱草属 Elymus | ||||||||
| 5鹅观草Elymus kamoji | p | 1.89 | ||||||
| 三 蔷薇科Rosaceae | ||||||||
| (六)委陵菜属Potentilla | ||||||||
| 6朝天委陵菜Potentilla supina | a/b | 3.18 | 16.55 | 4.22 | 1.47 | 10.8 | 8.31 | |
| 四 伞形科Apiaceae | ||||||||
| (七) 胡萝卜属Daucus | ||||||||
| 7野胡萝卜Daucus carota | b | 1.99 | 13.67 | 2.4 | 13.68 | |||
| 五 毛茛科Ranunculaceae | ||||||||
| (八)毛茛属 Ranunculus | ||||||||
| 8肉根毛茛Ranunculus polii | a | 5.98 | 14.13 | 7.87 | 1.32 | |||
| 六 蝶形花科Papilionaceae | ||||||||
| (九)黄芪属Astragalus | ||||||||
| 9紫云英Astragalus sinicus | b | 8.21 | 6.12 | 4.92 | 9.28 | |||
| 七 菊科Asteraceae | ||||||||
| (十) 蒿属 Artemisia | ||||||||
| 10艾Artemisia argyi | p | 4.61 | 10.85 | 2.13 | ||||
| 11蒌蒿Artemisia selengensis | p | 3.07 | ||||||
| (十一) 飞蓬属 Erigeron | ||||||||
| 12小蓬草Erigeron canadensis | a | 2.41 | 5.03 | 2.27 | 1.86 | |||
| (十二) 黄鹌菜属 Youngia | ||||||||
| 13黄鹌菜Youngia japonica | a | 1.43 | 3.58 | |||||
| (十三) 鼠曲草属Pseudognaphalium | ||||||||
| 14鼠曲草Pseudognaphalium affine | a | 3.25 | 1.57 | |||||
| (十四) 稻槎菜属 Lapsanastrum | ||||||||
| 15稻槎菜Lapsanastrum apogonoides | a | 4.59 | ||||||
| 八 茜草科Rubiaceae | ||||||||
| (十五)拉拉藤属 Galium | ||||||||
| 16六叶葎Galium hoffmeisteri | a | 4.25 | 7.48 | 3.55 | ||||
| 17四叶葎Galium bungei | p | 8.16 | 2.84 | 1.42 | ||||
| 九 旋花科Convolvulaceae | ||||||||
| (十六) 打碗花属 Calystegia | ||||||||
| 18打碗花Calystegia hederacea | a | 8.53 | ||||||
| 十 十字花科Brassicaceae | ||||||||
| (十七)蔊菜属 Rorippa | ||||||||
| 19两栖蔊菜Rorippa amphibia | p | 7.83 | ||||||
| (十八)碎米荠属 Cardamine | ||||||||
| 20弯曲碎米荠Cardamine flexuosa | a/b | 1.4 | 2.67 | |||||
| 十一 玄参科Scrophulariaceae | ||||||||
| (十九)通泉草属 Mazus | ||||||||
| 21通泉草Mazus japonicus | a | 1.42 | 4.81 | |||||
| 十二 蓼科Polygonaceae | ||||||||
| (二十)酸模属 Rumex | ||||||||
| 22齿果酸模Rumex dentatus | a | 3.15 | 2.4 | |||||
| 十三 牻牛儿苗科Geraniaceae | ||||||||
| (二十一)老鹳草属 Geranium | ||||||||
| 23野老鹳草Geranium carolinianum | a | 3.57 | 1.61 | |||||
| 十四 紫草科Boraginaceae | ||||||||
| (二十二)附地菜属 Trigonotis | ||||||||
| 24附地菜Trigonotis peduncularis | a/b | 3.55 | ||||||
| 十五 唇形科Lamiaceae | ||||||||
| (二十三)益母草属 Leonurus | ||||||||
| 25益母草Leonurus japonicus | a/b | 2.78 | ||||||
| (二十四)鼠尾草属 Salvia | ||||||||
| 26荔枝草Salvia plebeia | a/b | 1.55 | ||||||
2 The average number of species and diversity index of plant community in the marshland of Shengjin Lake升金湖洲滩植物群落样方物种平均数及多样性指数 |
| 时间 | 样方物种平均数/种 | Shannon-Wiener多样性指数 | |||||
| 低程区 | 中程区 | 高程区 | 低程区 | 中程区 | 高程区 | ||
| 2月 | 7.0 | 4.0 | 3.0 | 0.887 | 0.944 | 0.472 | |
| 4月 | 4.3 | 3.4 | 3.0 | 0.493 | 0.157 | 0.548 | |
| 时间 | Simpson多样性指数 | Pielou均匀度指数 | |||||
| 低程区 | 中程区 | 高程区 | 低程区 | 中程区 | 高程区 | ||
| 2月 | 0.411 | 0.498 | 0.261 | 0.442 | 0.656 | 0.433 | |
| 4月 | 0.238 | 0.069 | 0.309 | 0.334 | 0.244 | 0.497 | |
| 1 |
| 2 |
| 3 |
| 4 |
| 5 |
| 6 |
| 7 |
| 8 |
| 9 |
| 10 |
| 11 |
| 12 |
| 13 |
| 14 |
| 15 |
| 16 |
| 17 |
| 18 |
| 19 |
| 20 |
| 21 |
| 22 |
| 23 |
| 24 |
| 25 |
| 26 |
| 27 |
| 28 |
| 29 |
| 30 |
| 31 |
| 32 |
| 33 |
| 34 |
| 35 |
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}