
Received date: 2023-07-19
Revised date: 2023-10-24
Online published: 2026-03-12
Copyright
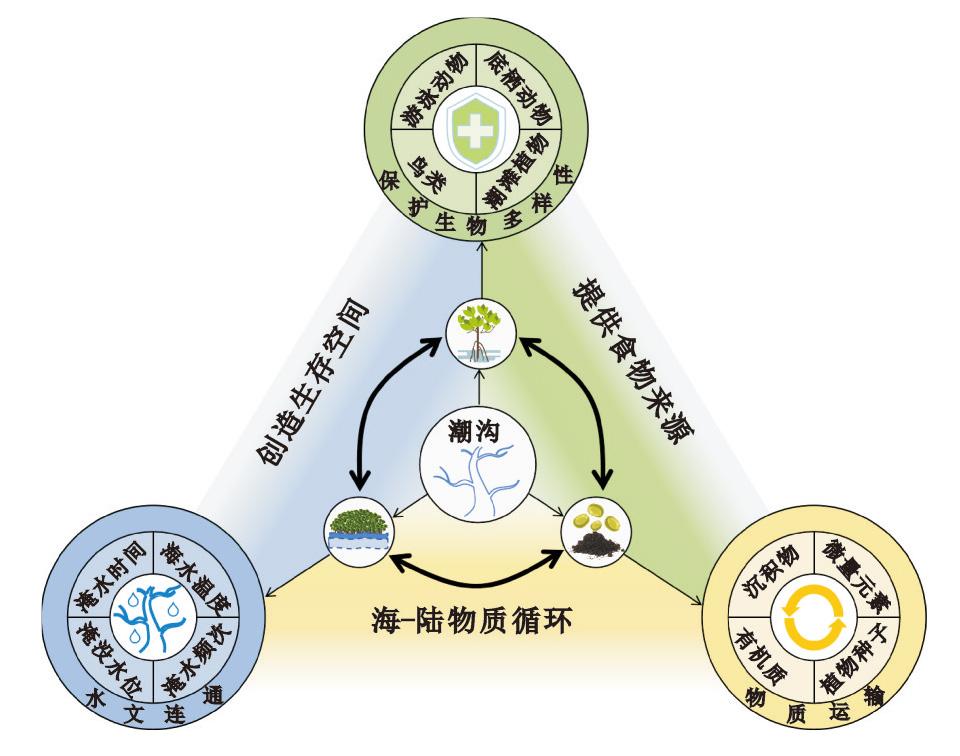
Tidal creeks, as unique components of coastal landform, have gradually attracted attention for their abilities of hydrological connectivity, material transportation and biodiversity. They can maintain the salt-and-water balance of tidal flats, transport sediments between ocean and land, be a spreading channel for plant seeds of coastal wetland, and provide habitats for marine organisms, which benefit the stability of coastal ecosystems. After extreme climate events and human activities, unchangeable shape and increasing biodiversity of tidal creeks is a signal of the restoration of coastal ecosystem. Therefore, it is of great significance to explore their morphological changes and ecological impact. Based on several researches of dynamic changes and ecological effects of tidal creeks in recent years, this paper reviews the classification methods, morphological characteristics, influencing factors and ecological effects of them, and puts forward three directions that need to be followed in the future: Spatial distribution and morphological characteristics of tidal creeks in different coastal wetlands with different influencing factors; Simulation and prediction of coastal seed transportation in tidal creeks. The relationships between the morphology, hydrological connectivity and ecological effect of tidal creeks.
AI Xihang , XIN Kun , ZENG Shujun . A Review on Evolution and Ecological Effects of Tidal Creeks[J]. Wetland Science, 2024 , 22(4) : 603 -616 . DOI: 10.13248/j.cnki.wetlandsci.2024.04.015
1 潮沟形态特征的概念及其计算公式Concepts and calculation formulas of tidal creek morphological characteristics |
| 潮沟形态特征 | 概念 | 计算公式 |
| 长度(Length) | 指研究区域内所有潮沟的总长 | L= ,其中,L为潮沟长度;ln为单条潮沟长度;n为潮沟数量 |
| 密度(Density) | 指单位面积潮滩上的潮沟总长度 | D= /A,其中,D为潮沟密度;A为潮滩面积 |
| 分汊率(Branchrate) | 指单位面积潮滩上潮沟分汊点的数量 | B=Nb/A,其中,B为潮沟分汊率;Nb为分汊点数量 |
| 曲率(Curvature) | 指潮沟实际长度与其两端点间的直线长度之比 | C=ln/dn,其中,C为潮沟曲率;dn为单条潮沟两端点间的直线距离 |
| 宽深比(W/D) | 潮沟宽度与深度的比值 | r=W/DP,其中,r为潮沟宽深比;W为潮沟宽度,DP为潮沟深度 |
| 分维值 (Fractal Dimension) | 以数学方法描述潮沟形态复杂程度 | FD=limr-0 ,其中,FD为潮沟分维值;r为包含整个潮沟的数个紧邻小矩形的边长,N(r)为包含潮沟所需的最少盒子数 |
| 1 |
| 2 |
| 3 |
| 4 |
| 5 |
| 6 |
| 7 |
| 8 |
| 9 |
| 10 |
| 11 |
| 12 |
| 13 |
| 14 |
| 15 |
| 16 |
| 17 |
| 18 |
| 19 |
| 20 |
| 21 |
| 22 |
| 23 |
| 24 |
| 25 |
| 26 |
| 27 |
| 28 |
| 29 |
| 30 |
| 31 |
| 32 |
| 33 |
| 34 |
| 35 |
| 36 |
| 37 |
| 38 |
| 39 |
| 40 |
| 41 |
| 42 |
| 43 |
| 44 |
| 45 |
| 46 |
| 47 |
| 48 |
| 49 |
| 50 |
| 51 |
| 52 |
| 53 |
| 54 |
| 55 |
| 56 |
| 57 |
| 58 |
| 59 |
| 60 |
| 61 |
| 62 |
| 63 |
| 64 |
| 65 |
| 66 |
| 67 |
| 68 |
| 69 |
| 70 |
| 71 |
| 72 |
| 73 |
| 74 |
| 75 |
| 76 |
| 77 |
| 78 |
| 79 |
| 80 |
| 81 |
| 82 |
| 83 |
| 84 |
| 85 |
| 86 |
| 87 |
| 88 |
| 89 |
| 90 |
| 91 |
| 92 |
| 93 |
| 94 |
| 95 |
| 96 |
| 97 |
| 98 |
| 99 |
| 100 |
| 101 |
| 102 |
| 103 |
| 104 |
| 105 |
| 106 |
| 107 |
| 108 |
| 109 |
| 110 |
| 111 |
| 112 |
| 113 |
| 114 |
| 115 |
| 116 |
| 117 |
| 118 |
| 119 |
| 120 |
| 121 |
| 122 |
| 123 |
| 124 |
| 125 |
| 126 |
| 127 |
| 128 |
| 129 |
| 130 |
| 131 |
| 132 |
| 133 |
| 134 |
| 135 |
| 136 |
| 137 |
| 138 |
| 139 |
| 140 |
| 141 |
| 142 |
| 143 |
| 144 |
| 145 |
| 146 |
| 147 |
| 148 |
| 149 |
| 150 |
| 151 |
| 152 |
| 153 |
| 154 |
| 155 |
| 156 |
| 157 |
| 158 |
| 159 |
| 160 |
| 161 |
| 162 |
| 163 |
| 164 |
| 165 |
| 166 |
| 167 |
| 168 |
| 169 |
| 170 |
| 171 |
| 172 |
| 173 |
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}