
Phosphate-solubilizing fungi: Isolation, characterization, and impact on soil as potential biofertilizers
Received date: 2025-08-07
Revised date: 2025-11-29
Accepted date: 2025-12-12
Online published: 2026-03-12
The escalating global demand for sustainable agriculture necessitates the development of effective biological alternatives to conventional chemical fertilizers, particularly those addressing phosphorus (P) use efficiency. This study focused on the isolation and detailed characterization of phosphate-solubilizing fungi from soil or compost to evaluate their impact and potential for use as biofertilizers. Fungal isolation was performed using serial dilution from various sources, followed by molecular and morphological characterization to identify promising strains. Four strains were ultimately selected and identified using morphological, biochemical, and molecular techniques: Aspergillus flavus (CM1), Penicillium crustosum (C3), Penicillium fellutanum (C4), and Metarhizium robertsii (J1). The most active strain was initially tested in liquid and solid media supplemented with synthetic P (Ca3(PO4)2) and was evaluated by measuring fungal biomass and P titration. This strain demonstrated good growth and activity, supporting an optimal temperature of 25°C, a pH of 3, an ammonium concentration of 1.5 g/L, and a glucose addition of 25.0 g/L. The biofertilization potential of the selected strains was then comprehensively evaluated through controlled experiments, including the optimization of growing conditions, quanti fication of soluble P under hermetic storage in soil, and measurement of soil fungal populations to assess their impact. P transformation experiments conducted in hermetic jars showed that CM1 had the highest CO2 release (approximately 7115.30 mg CO2/100 g soil) and the highest soluble P levels at the final sampling time (78.85 mg/L), thus outperforming the other strains. Furthermore, in soil hermetic jars, CM1 (reaching up to 26×104 CFU (colony forming units)/g soil) and C4 significantly enhanced soil microbial activity and P bioavailability. These results clearly highlight the potential of the selected fungal strains as biofertilizers to improve P availability and boost crop productivity in P-deficient soils.
Key words: phosphorus bioavailability; biofertilizers; fungi; soil; sustainable agriculture
Rim WERHENI AMMERI , Yasmine OCHI , Maroua OUESLETI , Hassen ABDENNACEUR , Najla SADFI ZOUAOUI . Phosphate-solubilizing fungi: Isolation, characterization, and impact on soil as potential biofertilizers[J]. Journal of Arid Land, 2026 , 18(2) : 339 -352 . DOI: 10.1016/j.jaridl.2026.02.007
Table 1 Selection of phosphate-solubilizing fungal strains in Pikovskaya (PVK) liquid and agar media supplemented with P at 25°C |
| Strain | Halo diameter (mm) | P content (mg/L) | Biomass of fungi (g) |
|---|---|---|---|
| J1 | 3.00±0.12 | 1.63±0.12 | 16.09±1.20 |
| C4 | 3.50±0.10 | 1.64±0.14 | 15.58±2.10 |
| CM1 | 4.00±0.01 | 1.62±0.21 | 20.92±2.30 |
| C3 | 2.50±0.20 | 1.61±0.36 | 11.68±1.65 |
Note: J1, Metarhizium robertsii; C4, Penicillium fellutanum; CM1, Aspergillus flavus; C3, Penicillium crustosum. Mean±SD; n=3. |
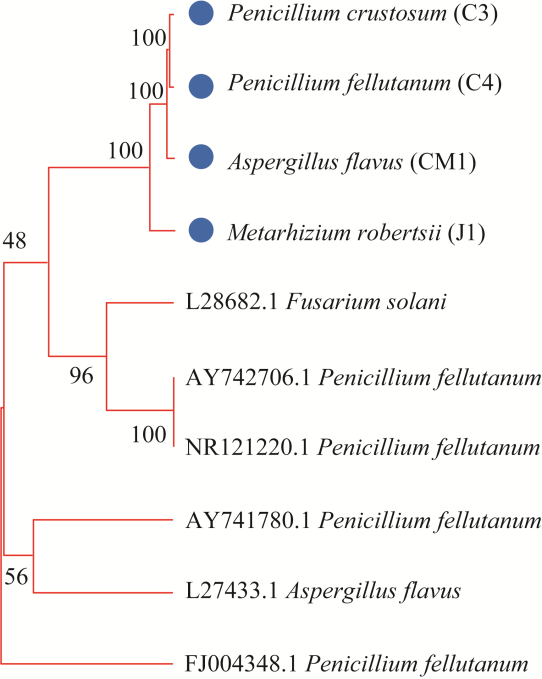
Fig. 1 Molecular evolutionary genetics analysis of fungal isolates. The number of 48, 56, 96, and 100 means bootstrap value, which indicates the strength of the evidence supporting the grouping of the isolates. |
Table 2 Effects of temperature, pH, glucose, and ammonium on phosphorus (P) and pellet weight formation for fungal strains in PVK liquid medium |
| Fungal strain | Temperature (°C) | pH | Glucose (g/L) | Ammonium (g/L) | |||||
|---|---|---|---|---|---|---|---|---|---|
| 25 | 35 | 3 | 8 | 5.0 | 25.0 | 1.5 | 2.5 | ||
| J1 | P (mg/L) | 152.10±12.00 | 300.72±12.30 | 8.64±2.30 | 9.52±1.02 | 73.48±1.36 | 55.89±2.30 | 11.75±1.01 | 7.25±0.98 |
| Pellet (g) | 17.68±8.20 | 8.39±1.20 | 140.32±12.30 | 133.20±12.20 | 14.95±1.50 | 13.23±1.50 | 4.79±0.89 | 8.34±1.20 | |
| CM1 | P (mg/L) | 75.55±3.20 | 267.90±14.30 | 7.65±0.80 | 1.67±0.45 | 43.11±2.35 | 20.37±3.10 | 10.29±1.60 | 7.42±0.95 |
| Pellet (g) | 6.52±2.00 | 7.49±0.90 | 117.00±14.30 | 15.00±1.30 | 9.23±0.78 | 7.44±0.65 | 51.75±11.02 | 3.44±0.47 | |
| C4 | P (mg/L) | 109.30±7.00 | 370.37±2.50 | 7.77±0.91 | 2.35±0.14 | 83.13±4.50 | 67.27±3.70 | 65.89±6.20 | 3.45±0.64 |
| Pellet (g) | 4.97±0.50 | 4.23±0.20 | 66.58±11.50 | 42.10±8.10 | 5.42±0.12 | 5.96±1.02 | 58.31±4.56 | 2.92±0.05 | |
| C3 | P (mg/L) | 80.03±1.20 | 310.30±10.20 | 7.21±1.20 | 7.61±0.98 | 90.72±5.20 | 130.37±15.10 | 43.48±12.30 | 4.41±0.01 |
| Pellet (g) | 6.48±1.30 | 10.74±1.20 | 71.20±2.50 | 88.32±11.04 | 13.20±0.47 | 6.48±0.69 | 76.24±14.20 | 2.59±0.34 | |
Note: Mean±SD; n=3. |
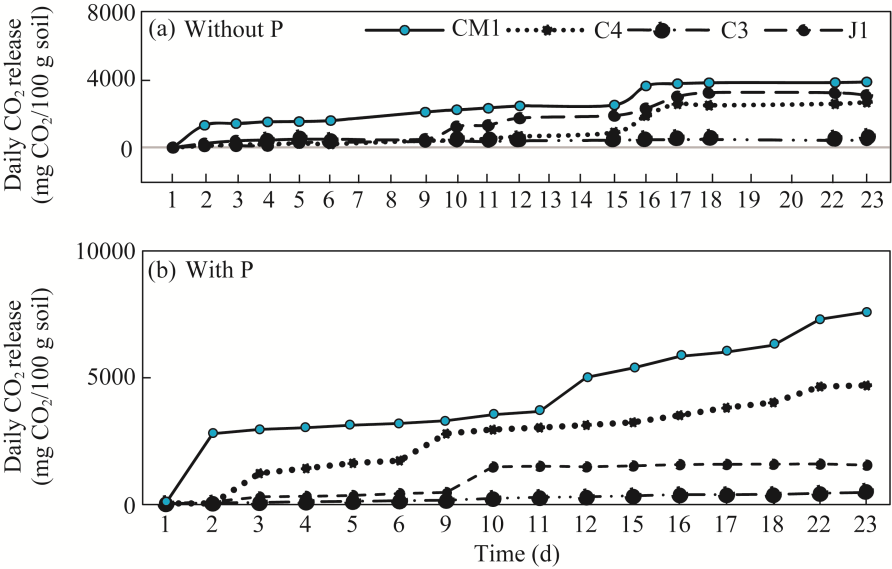
Fig. 2 Daily CO2 release from soil inoculated with fungal strains C4, C3, J1, and CM1, without (a) or with (b) phosphorus (P) addition. CM1, Aspergillus flavus; C4, Penicillium fellutanum; J1, Metarhizium robertsii; C3, Penicillium crustosum. |
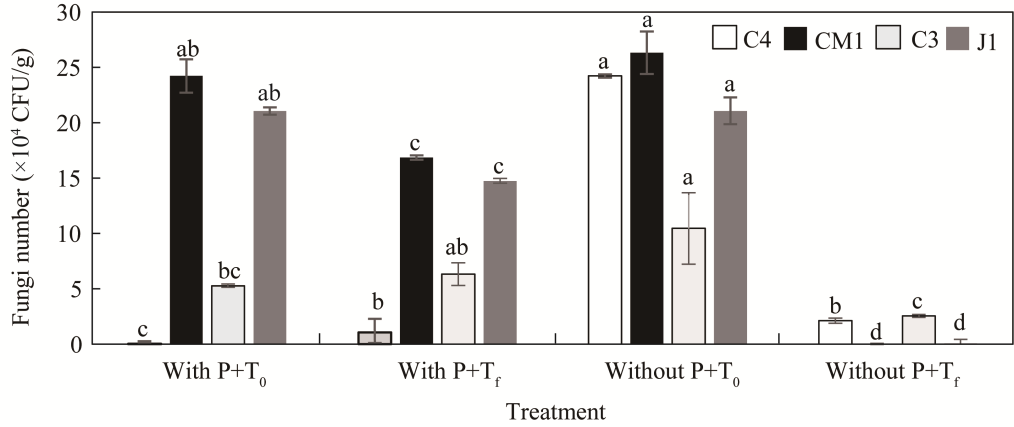
Fig. 3 Effect of P on fungal abundance in soil inoculated with strains C4, C3, J1, and CM1 with or without P. T0, initial sampling time, Tf, final sampling time; CFU, colony forming units. Different lowercase letters within the same treatment indicate significant differences among different strains at P<0.05 level. Bars are standard deviations. |
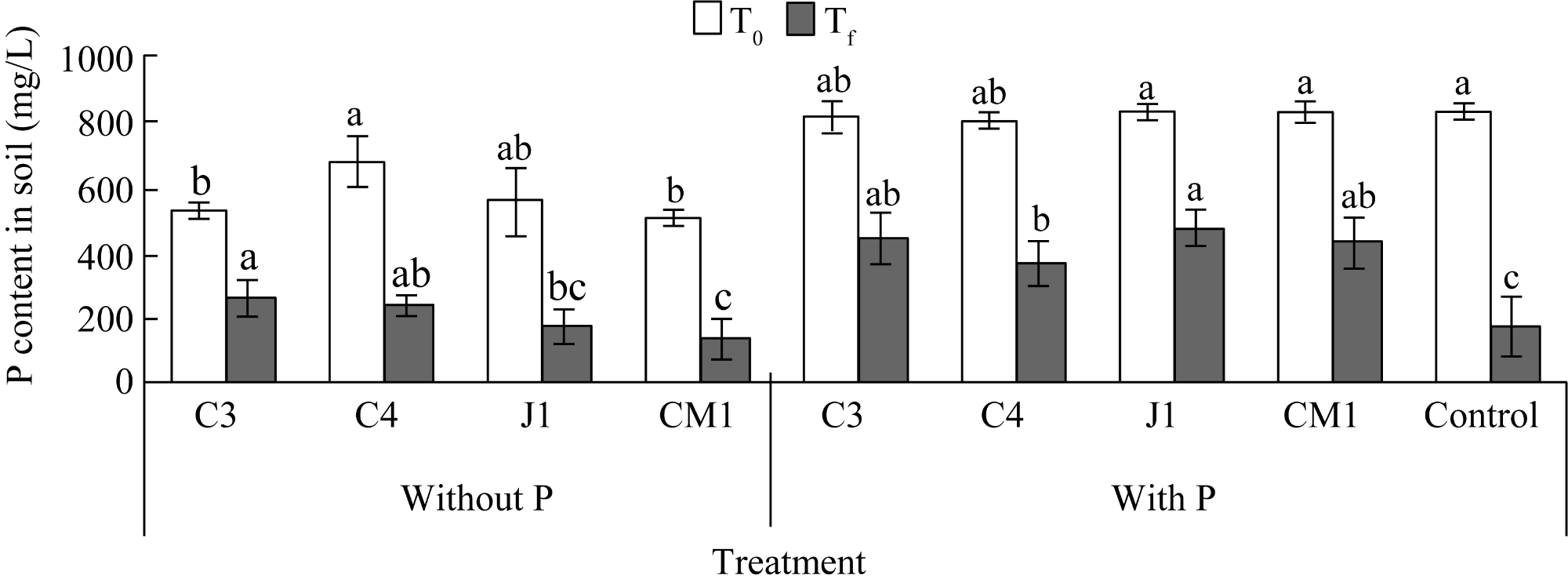
Fig. 4 Variations in P content in soil amended with Ca3(PO4)2 under different treatments. Different lowercase letters within the same addition and the same sapling time indicate significant differences among different treatments at P<0.05 level. Bars are standard deviations. |
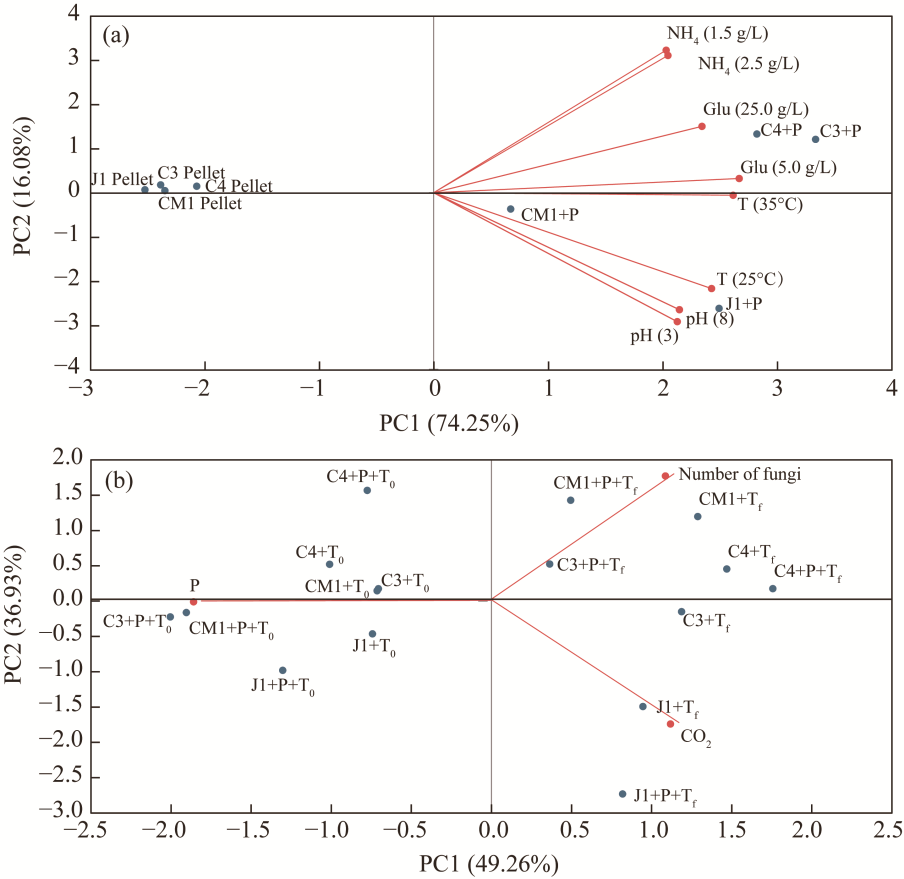
Fig. 5 Principal component analysis (PCA) of fungal responses under different experimental conditions. (a), liquid medium; (b), soil condition. PC, principal component; T, temperature; Glu, glucose; NH4, ammonium sulfate. |
| [1] |
|
| [2] |
APHA (American Public Health Association). 1998. Standard Methods for the Examination of Water and Wastewater (20th ed.). Washington D. C.: American Public Health Association.
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}