
Flammability of plant communities in arid and semi-arid ecosystems: Identifying key drivers and management implications
Received date: 2025-07-12
Revised date: 2025-11-01
Accepted date: 2025-11-21
Online published: 2026-03-12
Arid and semi-arid ecosystems are prone to extensive fires due to specific climatic conditions, sparse vegetation cover, and high density of fine fuels. Understanding the flammability characteristics of land covers is essential for fire management and designing land restoration programs in arid and semi-arid ecosystems. This study provided a new approach to evaluate the flammability of shrublands and woodlands using flammability indices (FIs) including time to ignition (TI), duration of combustion (DC), and flame height (FH) of plant species and their relative frequencies in the Dalfard Basin of southeastern Iran. The results showed that there was a significant difference in FIs between land covers. Shrublands had higher flammability potential compared with woodlands. Plant moisture content had a negative relationship with TI (P<0.010) and no significant relationship with DC and FH (P>0.050). Artemisia spp., Astragalus gossypinus Fischer, Amygdalus scoparia Spach, and Cymbopogon jwarancusa (Jones) Schult. had the highest FI. Tree species such as Rhazya stricta Decne., and Pistacia atlantica Desf. showed greater resistance to fire. Using principal component analysis, the relationship between species and FIs was examined, and TI of wet fuel was the most important FI in relation to species. Structural equation model showed that life form (P<0.001) was the most important flammability driver. Precipitation (P<0.010) and legume species (P<0.010) were significantly related to the flammability in arid land. This study emphasizes the importance of managing high-risk species and using resistant species in vegetation restoration and shows that combining species FIs with their abundance is an effective tool for assessing fire risk and fuel management at the plant community scale.
Mosayeb HOJATI , Azam KHHOSRAVI MASHIZI . Flammability of plant communities in arid and semi-arid ecosystems: Identifying key drivers and management implications[J]. Journal of Arid Land, 2026 , 18(2) : 304 -321 . DOI: 10.1016/j.jaridl.2026.02.005
Table 1 Environmental and plant characters in the Dalfard Basin, Iran |
| Plant type | Abbreviation | Elevation (m) | Precipitation (mm) | Temperature (°C) | |
|---|---|---|---|---|---|
| Minimum | Maximum | ||||
| Artemisia aucheri Boiss_Astragalus gossypinus Fischer | Ar_As | 2750 | 3672 | 450 | 12 |
| Pistacia atlantica Desf._A. gossypinus | Pi_As | 2405 | 3500 | 450 | 14 |
| Amygdalus scoparia L._A. gossypinus | Am_As | 2150 | 2385 | 360 | 18 |
| A. scoparia_Ebenus stellate Boiss. | Am_Eb | 1920 | 2200 | 350 | 18 |
| A. scoparia_Cymbopogon jwarancusa (Jones) Schult. | Am_Cy | 1620 | 1840 | 260 | 22 |
| Tecomella undulata (Sm.) Seem | Te | 1230 | 1850 | 250 | 22 |
| Rhazya stricta Decne. | Rh | 633 | 1220 | 150 | 34 |
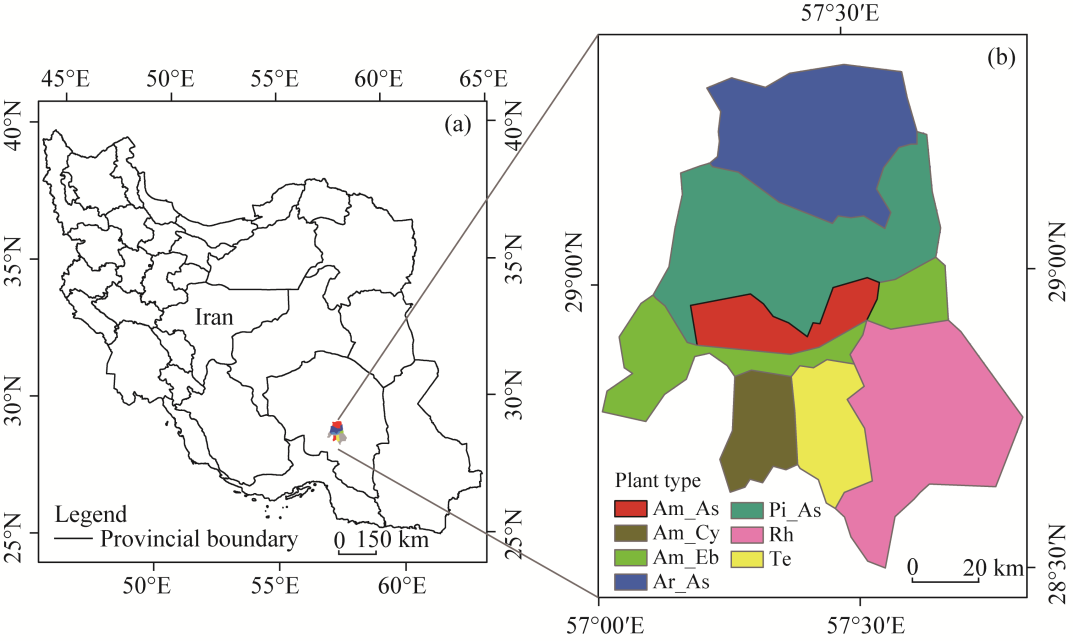
Fig. 1 Study area (a) and distribution of plant types in the Dalfard Basin, Iran (b). Am_As, Amygdalus scoparia L._A. gossypinus; Am_Cy, A. scoparia_Cymbopogon jwarancusa (Jones) Schult.; Am_Eb, A. scoparia_Ebenus stellate Boiss.; Ar_As, Artemisia aucheri Boiss_Astragalus gossypinus Fischer; Pi_As, Pistacia atlantica Desf._A. gossypinus; Rh, Rhazya stricta Decne.; Te, Tecomella undulata (Sm.) Seem. |
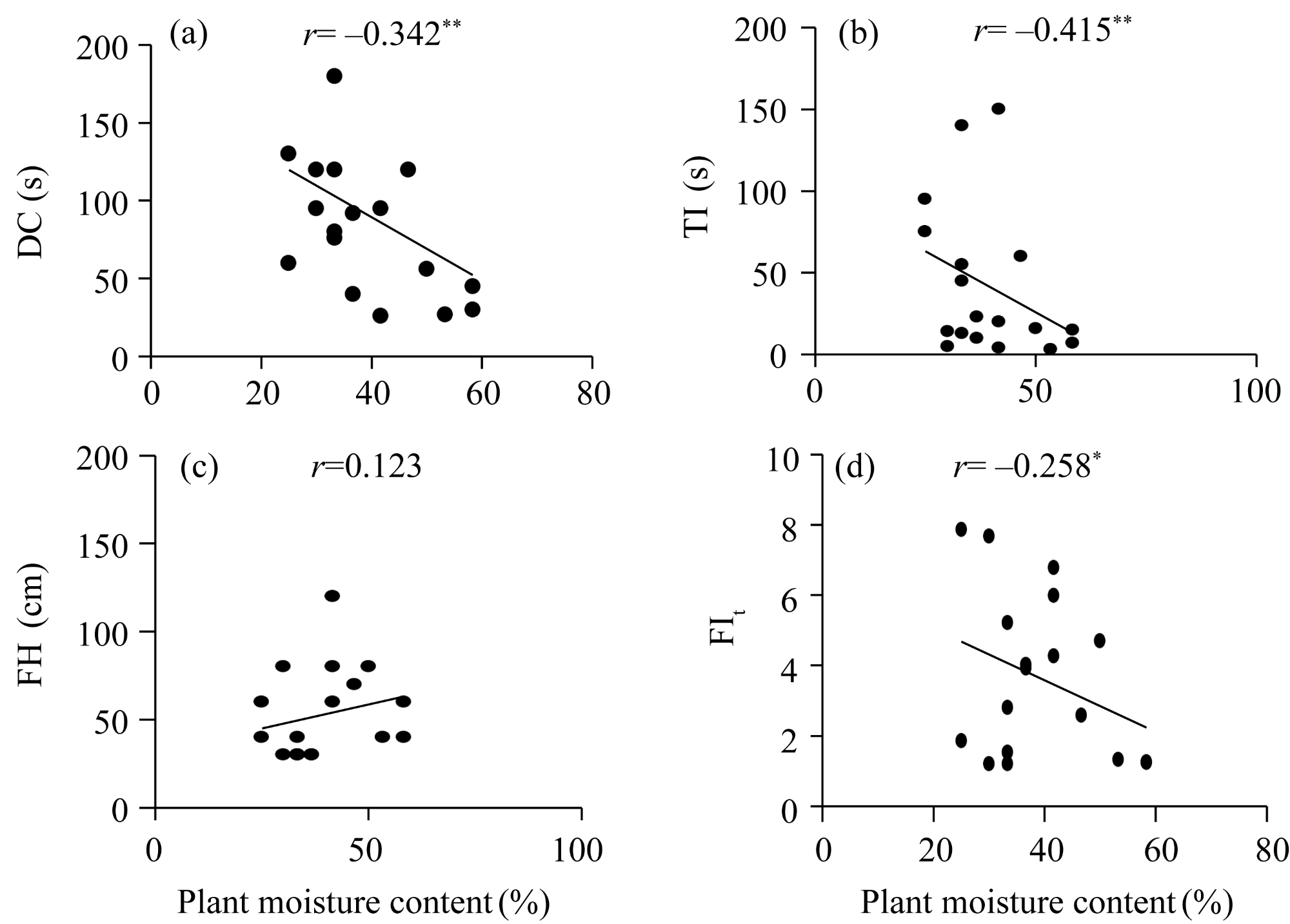
Fig. 2 Relationship between plant moisture content and flammability indices (FIs). (a), DC (duration of combustion); (b), TI (time to ignition); (d), FH (flame height); (e), FIt (FI of plant community). *, P<0.050 level; **, P<0.010 level. |
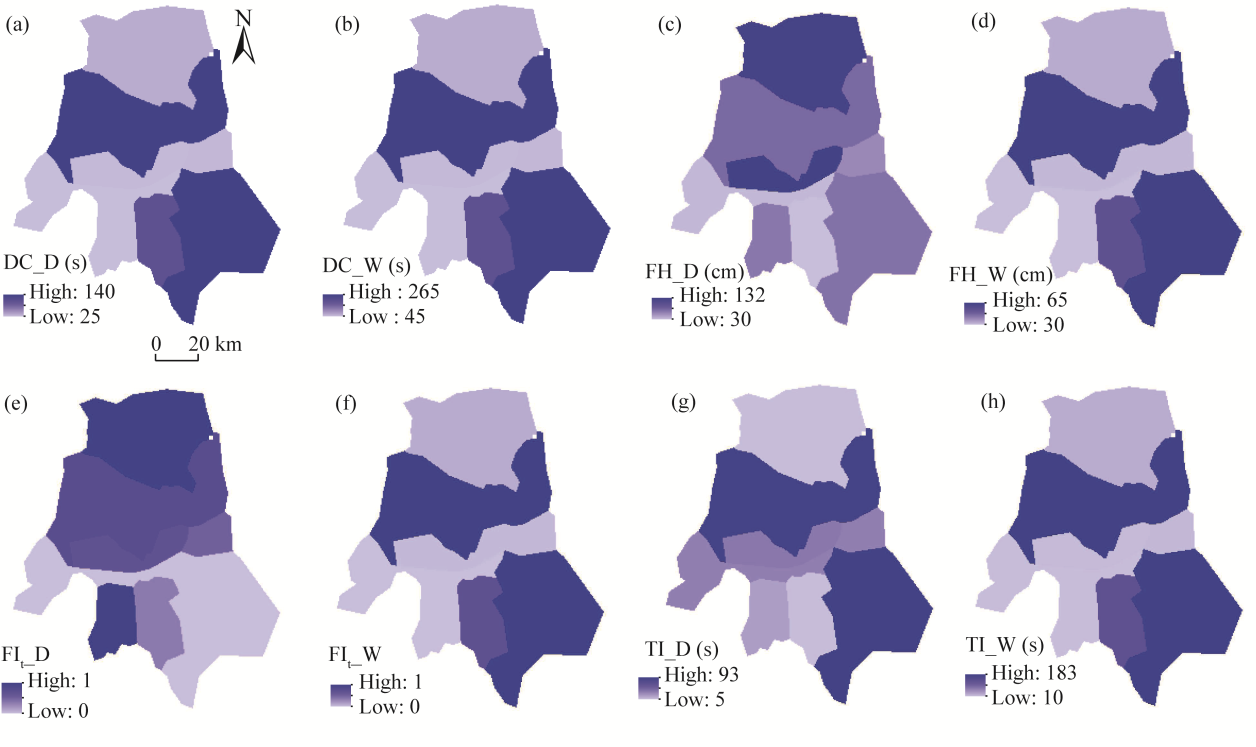
Fig. 3 Spatial distribution of FIs in the Dalfard Basin, Iran. (a), DC_D (dry fuel); (b), DC_W (wet fuel); (c), FH_D; (d), FH_W; (e), FIt_D; (f), FIt _W; (g), TI_D; (h), TI_W. |
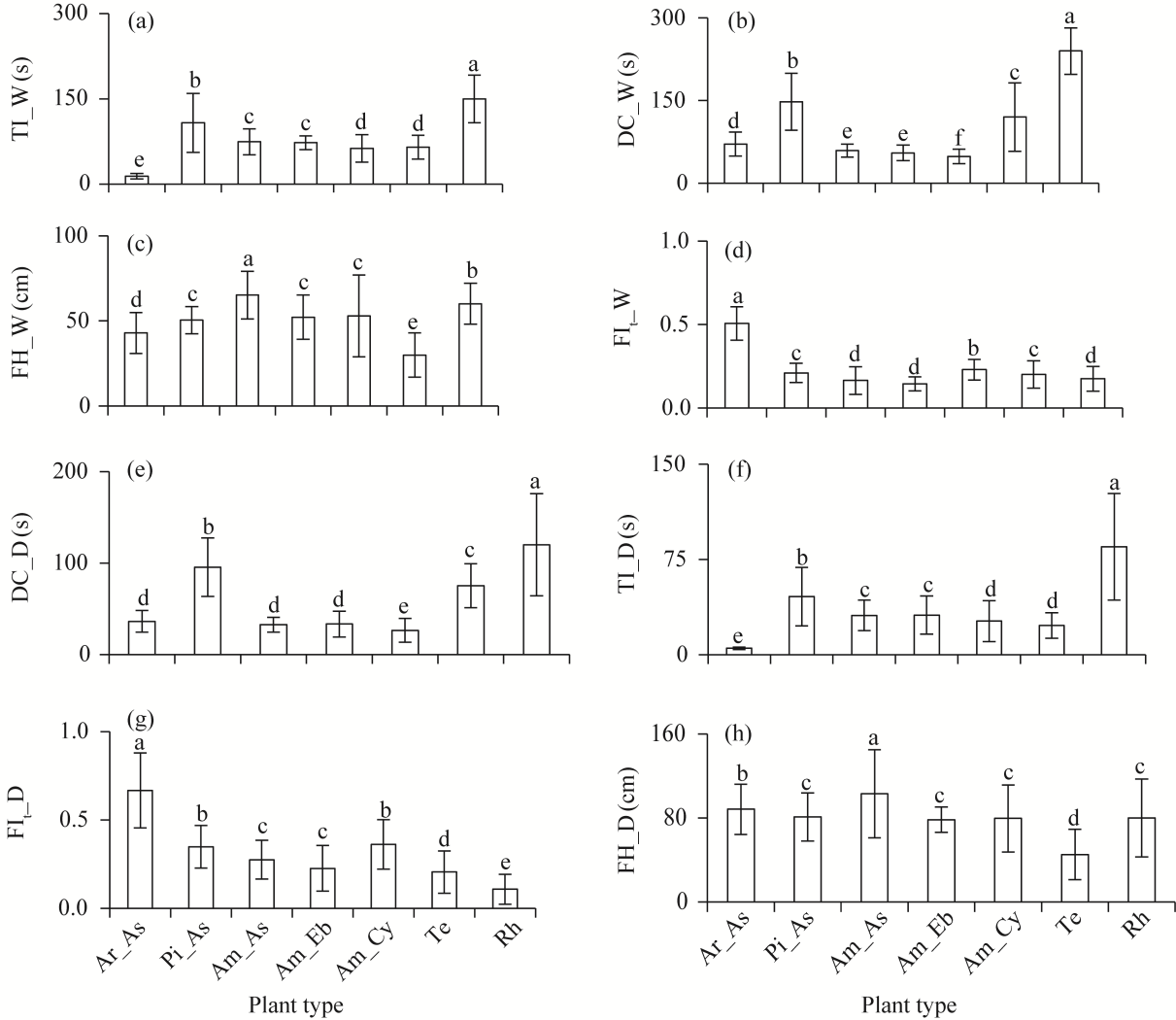
Fig. 4 Comparison of FIs among different plant types. (a), TI_W; (b), DC_W; (c), FH_W; (d), FIt_W; (e), DC_D; (f), TI_D; (g), FIt_D; (h), FH_D. Different lowercase letters indicate significant differences among plant types at P<0.050 level. Bars are standard errors. |
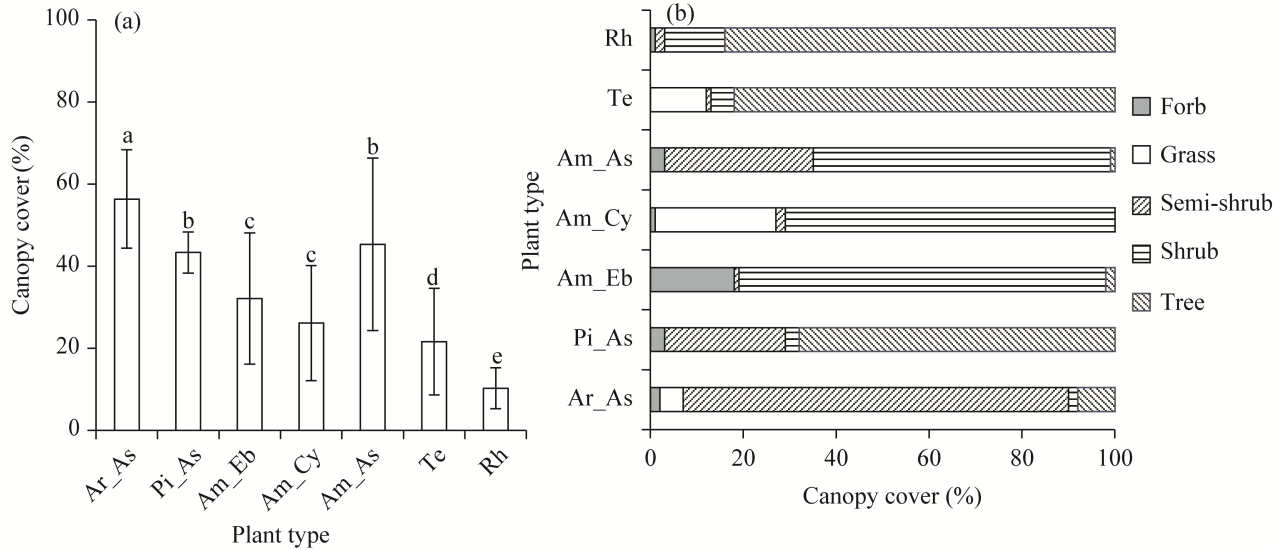
Fig. 5 Comparison of canopy cover in different plant types (a) and different life forms of plants (b). Different lowercase letters in Figure 5a indicate significant differences among different plant types at P<0.050 level. Bars are standard errors. |
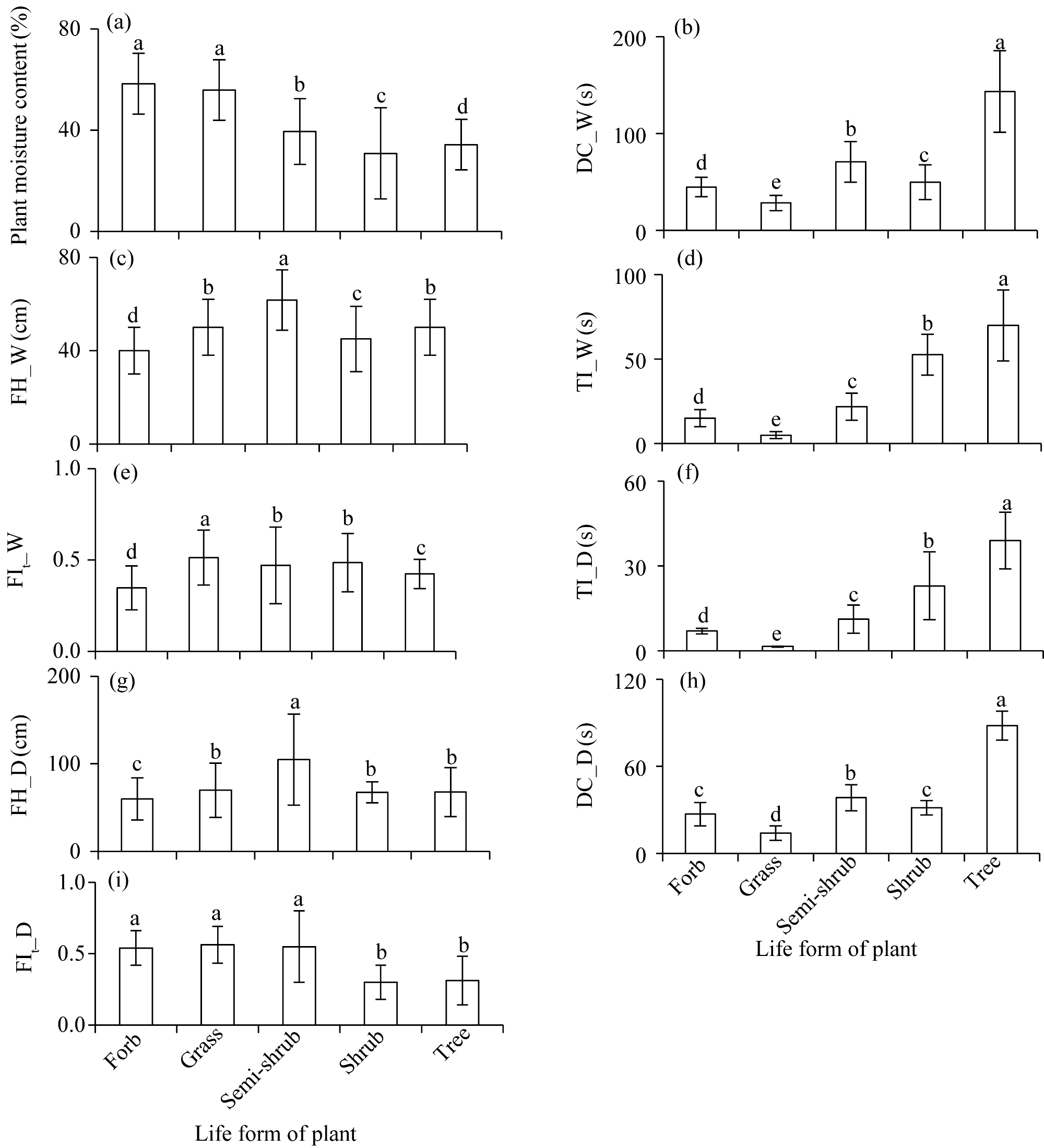
Fig. 6 Comparison of FIs among different life forms of plants. (a), plant moisture content; (b), DC_W; (c), FH_W; (d), TI_W; (e), FIt_W; (f), TI_D; (g), FH_D; (h), DC_D; (i), FIt_D. Different lowercase letters indicate significant differences among different life forms of plants at P<0.050 level. Bars are standard errors. |
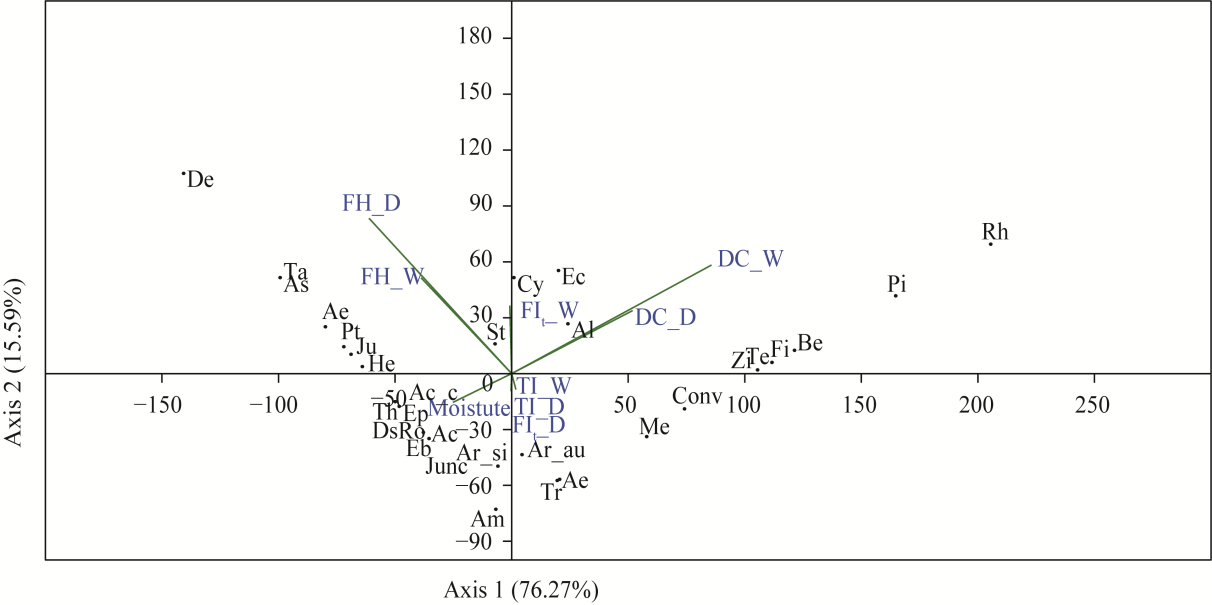
Fig. 7 Distribution of plant species in relation to FI based on principal component analysis (PCA). Ar_au, Artemisia aucheri Boiss; Ar_si, Artemisia sieberi Besser; As, Astragalus gossypinus Fischer; Pi, Pistacia atlantica Desf.; Am, Amygdalus scoparia L.; Cy, Cymbopogon jwarancusa (Jones) Schult.; Conv, Convolvulus arvensis L.; Ds, Descurainia sophia (L.) Webb ex Prantl; Eb, Ebenus stellata Boiss.; Pt, Pteropyrum aucheri Jaub.&Spach subsp. aucheri; Ro, Roemeria argemone (L.) C. Morales R. Mend.&Romero García; Ac, Acantholimon glandulosum Hand.-Mazz.; Tr, Trifolium pratense L. subsp. pretense; Ec, Echinops aucheri Boiss.; Me, Medicago sativa L.; Al, Allium jesdianum Boiss.&Buhse.; Fi, Ficus carica subsp. rupestris (Boiss.) Browicz; Ep, Ephedra ciliata Fisch.&C. A. Mey.; St, Stipa haussknechtii Boiss.; Junc, Juncus inflexus L.; Ae, Aerva javanica (Burm. f.) Juss.; Ac_c, Acantholimon chlorostegium Rech. f.&Schiman-Czeika; Be, Berberis integerrima Bunge; Ta, Tamarix androssowii Litv.; Zi, Ziziphus spina-christi (L.) Desf. Ju, Juniperus excelsa M.Bieb.; He, Heliotropium aucheri DC.; De, Descurainia sophia (L.) Webb ex Prantl. Th, Thymus daenensis subsp. lancifolius (Celak.) Jalas; Co, Cornulaca monacantha Delile. |
Table 2 Principal component analysis (PCA) loading for flammability indices (FIs) using the first two axes |
| Index | Axis 1 | Axis 2 | Index | Axis 1 | Axis 2 |
|---|---|---|---|---|---|
| Plant moisture | -0.231 | -0.124 | TI_D | 0.123 | 0.062 |
| DC_W | 0.454 | 0.245 | FH_D | -0.324 | 0.134 |
| TI_W | 0.524 | 0.214 | FIt_D | 0.128 | 0.051 |
| FH_W | -0.312 | 0.103 | Eigenvalue | 2.330 | 1.030 |
| FIt_W | 0.131 | 0.052 | Variance (%) | 76.27 | 15.59 |
| DC_D | 0.115 | 0.024 |
Note: The significant value is shown in bold. DC, duration of combustion; TI, time to ignition; FH, flame height; FIt, FI of plant community; D, dry fuel; W, wet fuel. |
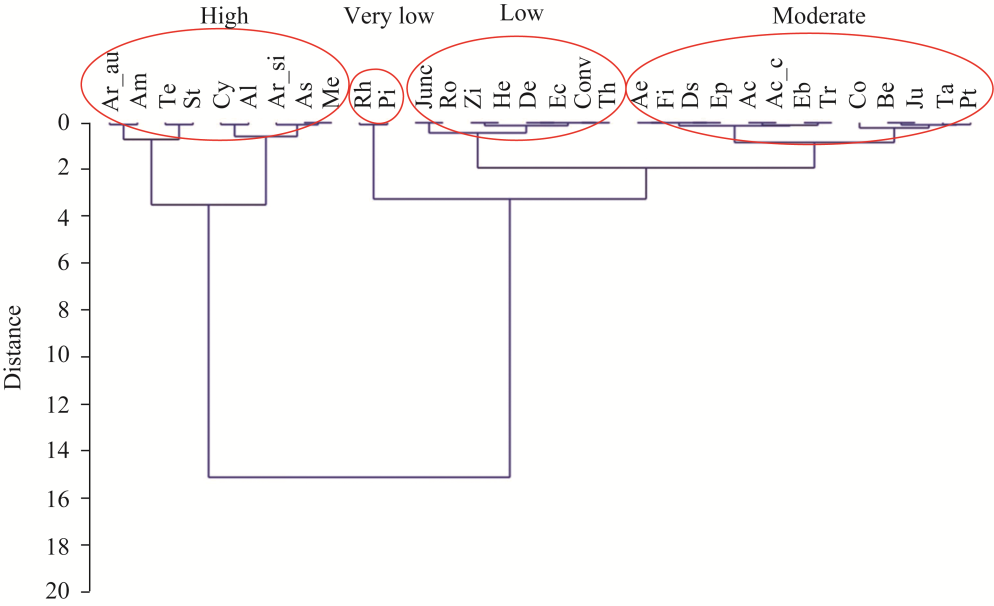
Fig. 8 Clustering plant species in very low, low, moderate, and high classes of FIs |
Table 3 Average variance extracted (AVE), composite reliability (CR), and total impacts of plant traits and environmental factors related to flammability using structural equation model (SEM) |
| Variable | AVE | CR | Impact |
|---|---|---|---|
| Canopy size | 0.754 | 0.836 | 0.276* |
| Species height | 0.824 | 0.875 | 0.062 |
| Legume species | 0.817 | 0.924 | 0.375** |
| C4 species | 0.819 | 0.832 | 0.369** |
| Leaf life span | 0.852 | 0.925 | -0.296* |
| Life form | 0.832 | 0.872 | -0.487*** |
| Temperature | 0.732 | 0.824 | 0.152* |
| Precipitation | 0.933 | 0.941 | 0.389** |
| Species family | 0.795 | 0.825 | 0.241* |
| Elevation | 0.758 | 0.865 | 0.162* |
| Canopy cover | 0.826 | 0.924 | 0.173* |
| Species diversity | 0.791 | 0.837 | 0.190* |
| Flammability | 0.975 | 0.982 | - |
Note: *, P<0.050 level; **, P<0.010 level; ***, P<0.001 level. ''-'' indicates no value. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}