
Wind and sand control in composite shelterbelts combining Cyperus esculentus with trees and shrubs: Evidence from wind tunnel and field studies
Received date: 2025-05-06
Revised date: 2025-08-19
Accepted date: 2025-08-25
Online published: 2026-03-12
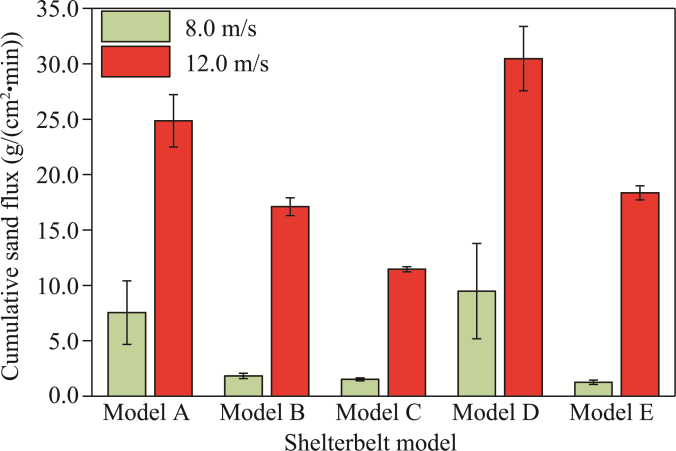
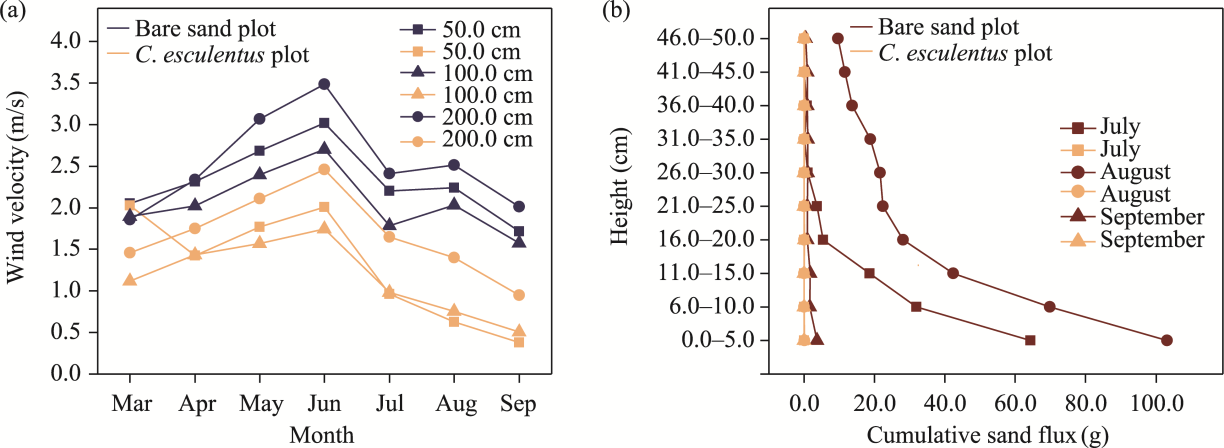
Cyperus esculentus (C. esculentus), a desert-adapted plant species with both ecological and economic value, has been widely cultivated in northern China's sandy regions. However, limited studies have investigated the performance of composite shelterbelts that integrate C. esculentus. This study systematically evaluated five shelterbelt models—Populus euphratica (P. euphratica), P. euphratica-C. esculentus composite, P. euphratica-nylon net-C. esculentus composite, Tamarix chinensis (T. chinensis), and T. chinensis-C. esculentus composite—using wind tunnel experiments and field observations. Sediment flux was measured at a normalized downwind distance (x/h) of 5, where x refers to the distance from the front edge (upwind side) of the shelterbelt for upwind measurements, and the distance from the rear edge (downwind side) for downwind measurements, and h represents the canopy height. Wind velocity was measured at x/h of -2, -1, 1, 2, 3, 5, and 7, and sand flux was measured at x/h=5, under initial wind velocities of 8.0 and 12.0 m/s. The results indicated that the P. euphratica-nylon net-C. esculentus composite was the most effective in reducing wind velocity, followed by the P. euphratica-C. esculentus composite. In contrast, the P. euphratica and T. chinensis exhibited relatively weaker wind reduction capabilities. Regarding sand flux, under moderate wind velocity (8.0 m/s), both the P. euphratica-C. esculentus composite and P. euphratica-nylon net-C. esculentus composite demonstrated the lowest sand flux values. However, under high wind velocity (12.0 m/s), the P. euphratica-nylon net-C. esculentus composite significantly outperformed the other shelterbelt models in sand retention, highlighting its superior windbreak and sand fixation efficacy. Field observations further validated the windbreak and sand fixation effects of C. esculentus. Comparisons between the bare sand plot and C. esculentus plot within protective forests demonstrated that planting C. esculentus can provide substantial ecological benefits in windbreak and sand-fixation. These findings, reinforced by field observations, strengthen the wind tunnel experiment results and highlight the critical role of C. esculentus in enhancing the performance of composite shelterbelts for desert ecological restoration.
Key words: Cyperus esculentus; composite shelterbelt; wind tunnel; sand flux; desert ecosystems
NIE Bixia , SHEN Xin , LIU Yalan , LI Xiangyi . Wind and sand control in composite shelterbelts combining Cyperus esculentus with trees and shrubs: Evidence from wind tunnel and field studies[J]. Journal of Arid Land, 2026 , 18(2) : 263 -279 . DOI: 10.1016/j.jaridl.2026.02.004
Fig. 1 Customized plastic model pictures of Populus euphratica (a), Tamarix chinensis (b), and Cyperus esculentus (c), as well as comparison among three plant species (d) |
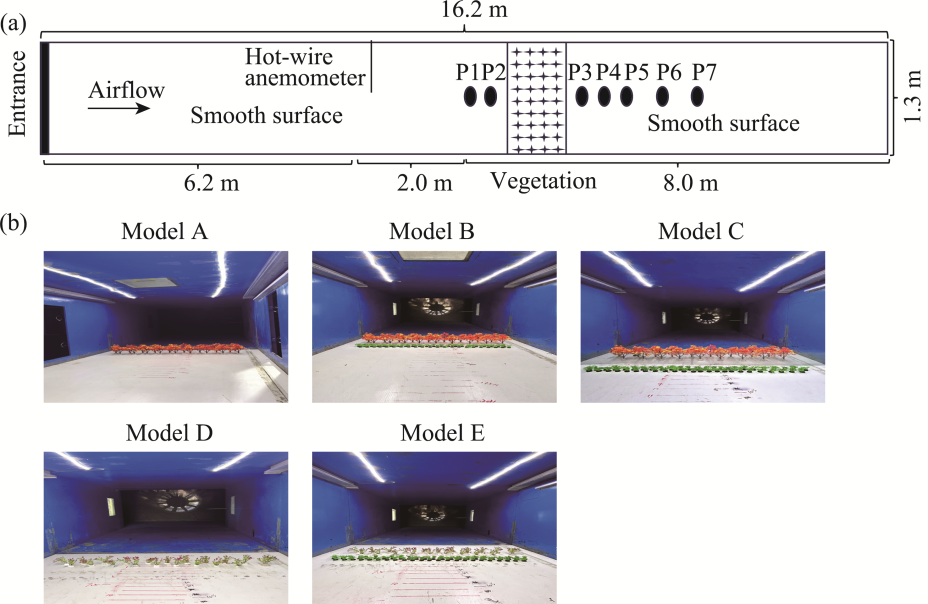
Table 1 Pitot tube locations of model A (Populus euphratica) in the wind tunnel |
| Pitot tube | Location | x (cm) | x/h |
|---|---|---|---|
| P1 | 40.0 cm upwind of the vegetated surface (820.0 cm downwind from the leading edge of the entrance of work section) | -40.0 | -2 |
| P2 | 20.0 cm upwind of the vegetated surface (840.0 cm downwind from the leading edge of the entrance of work section) | -20.0 | -1 |
| P3 | 30.0 cm downwind from the leading edge of vegetation (890.0 cm downwind of the entrance of work section ) | 20.0 | 1 |
| P4 | 50.0 cm downwind from the leading edge of vegetation (910.0 cm downwind of the entrance of work section ) | 40.0 | 2 |
| P5 | 70.0 cm downwind from the leading edge of vegetation (930.0 cm downwind of the entrance of work section ) | 60.0 | 3 |
| P6 | 110.0 cm downwind from the leading edge of vegetation (970.0 cm downwind of the entrance of work section ) | 100.0 | 5 |
| P7 | 150.0 cm downwind from the leading edge of vegetation (1010.0 cm downwind of the entrance of work section ) | 140.0 | 7 |
Note: x/h is the normalized downwind distance; x refers to the distance from the front edge (upwind side) of the shelterbelt for upwind measurements, or the distance from the rear edge (downwind side) for downwind measurements; h represents the canopy height. |
Table 2 Pitot tube locations of models B (P. euphratica-Cyperus esculentus composite) and C (P. euphratica-nylon net-C. esculentus composite) in the wind tunnel |
| Pitot tube | Location | x (cm) | x/h |
|---|---|---|---|
| P1 | 40.0 cm upwind of the vegetated surface (820.0 cm downwind from the leading edge of the entrance of work section) | 40.0 | -2 |
| P2 | 20.0 cm upwind of the vegetated surface (840.0 cm downwind from the leading edge of the entrance of work section) | 20.0 | -1 |
| P3 | 42.0 cm downwind from the leading edge of vegetation (902.0cm downwind of the entrance of work section) | 20.0 | 1 |
| P4 | 62.0 cm downwind from the leading edge of vegetation (922.0 cm downwind of the entrance of work section) | 40.0 | 2 |
| P5 | 82.0 cm downwind from the leading edge of vegetation (942.0 cm downwind of the entrance of work section) | 60.0 | 3 |
| P6 | 122.0 cm downwind from the leading edge of vegetation (982.0 cm downwind of the entrance of work section) | 100.0 | 5 |
| P7 | 162.0 cm downwind from the leading edge of vegetation (1022.0 cm downwind of the entrance of work section) | 140.0 | 7 |
Table 3 Pitot tube locations of model D (Tamarix chinensis) in the wind tunnel |
| Pitot tube | Location | x (cm) | x/h |
|---|---|---|---|
| P1 | 20.0 cm upwind of the vegetated surface (820 cm downwind from the leading edge of the entrance of work section) | -20.0 | -2 |
| P2 | 10.0 cm upwind of the vegetated surface (830 cm downwind from the leading edge of the entrance of work section) | -10.0 | -1 |
| P3 | 15.0 cm downwind from the leading edge of vegetation (855.0 cm downwind of the entrance of work section) | 10.0 | 1 |
| P4 | 25.0 cm downwind from the leading edge of vegetation (865.0 cm downwind of the entrance of work section) | 20.0 | 2 |
| P5 | 35.0 cm downwind from the leading edge of vegetation (875.0 cm downwind of the entrance of work section) | 30.0 | 3 |
| P6 | 55.0 cm downwind from the leading edge of vegetation (895.0 cm downwind of the entrance of work section) | 50.0 | 5 |
| P7 | 75.0 cm downwind from the leading edge of vegetation (915.0 cm downwind of the entrance of work section) | 70.0 | 7 |
Table 4 Pitot tube locations of model E (T. chinensis-C. esculentus composite) in the wind tunnel |
| Pitot tube | Location | x (cm) | x/h |
|---|---|---|---|
| P1 | 20.0 cm upwind of the vegetated surface (820.0 cm downwind from the leading edge of the entrance of work section) | -20.0 | -2 |
| P2 | 10.0 cm upwind of the vegetated surface (830.0 cm downwind from the leading edge of the entrance of work section) | -10.0 | -1 |
| P3 | 22.0 cm downwind from the leading edge of vegetation (862.0 cm downwind of the entrance of work section) | 10.0 | 1 |
| P4 | 32.0 cm downwind from the leading edge of vegetation (872.0 cm downwind of the entrance of work section) | 20.0 | 2 |
| P5 | 42.0 cm downwind from the leading edge of vegetation (882.0 cm downwind of the entrance of work section) | 30.0 | 3 |
| P6 | 62.0 cm downwind from the leading edge of vegetation (902.0 cm downwind of the entrance of work section) | 50.0 | 5 |
| P7 | 82.0 cm downwind from the leading edge of vegetation (922.0 cm downwind of the entrance of work section) | 70.0 | 7 |
Fig. 2 Schematic diagram of the Pitot tube system used for wind velocity measurements in the wind tunnel (a) and wind velocity sampling process for the five shelterbelt models A-E during wind tunnel experiments (b). P1-P7 indicate seven sequential sampling positions where wind velocity was measured by a single Pitot tube. |
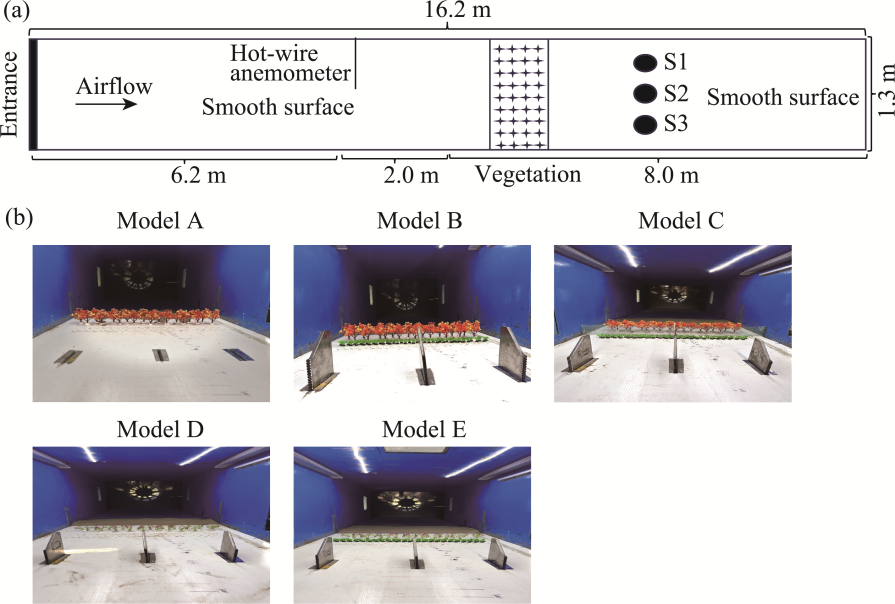
Fig. 3 Schematic diagram of the sand trap system used in the wind tunnel working section (a) and sand sampling process for the five shelterbelt models A-E during wind tunnel experiments (b). S1-S3 indicate three replicated sand traps. |
Table 5 Location of sand samplers in the wind tunnel |
| Model | Location | x (cm) | x/h |
|---|---|---|---|
| Model A | 110.0 cm downwind from the leading edge of vegetation (970.0 cm downwind of the entrance of work section ) | 100.0 | 5 |
| Model B | 122.0 cm downwind from the leading edge of vegetation (982.0 cm downwind of the entrance of work section) | 100.0 | 5 |
| Model C | 15.0 cm downwind from the leading edge of vegetation (855.0 cm downwind of the entrance of work section) | 100.0 | 5 |
| Model D | 55.0 cm downwind from the leading edge of vegetation (895.0 cm downwind of the entrance of work section) | 50.0 | 5 |
| Model E | 62.0 cm downwind from the leading edge of vegetation (902.0 cm downwind of the entrance of work section) | 50.0 | 5 |
Fig. 4 Photographs showing the bare sand plot (a) and C. esculentus plot (b), along with the experimental layout diagram (c) |
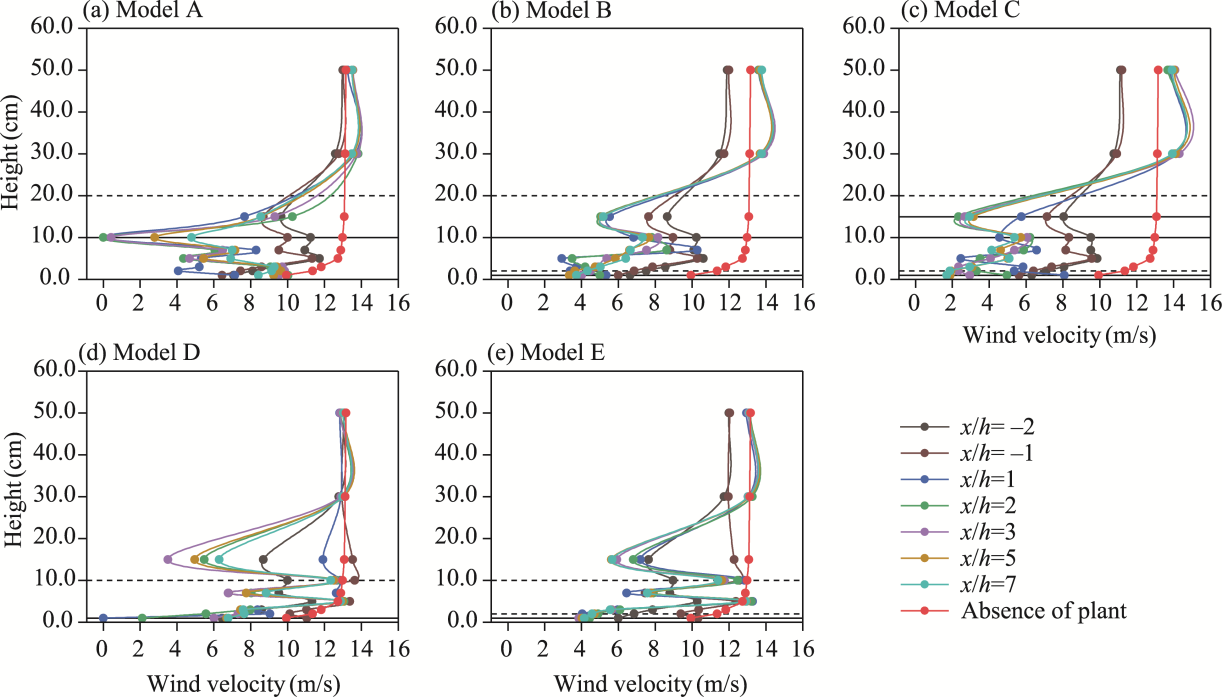
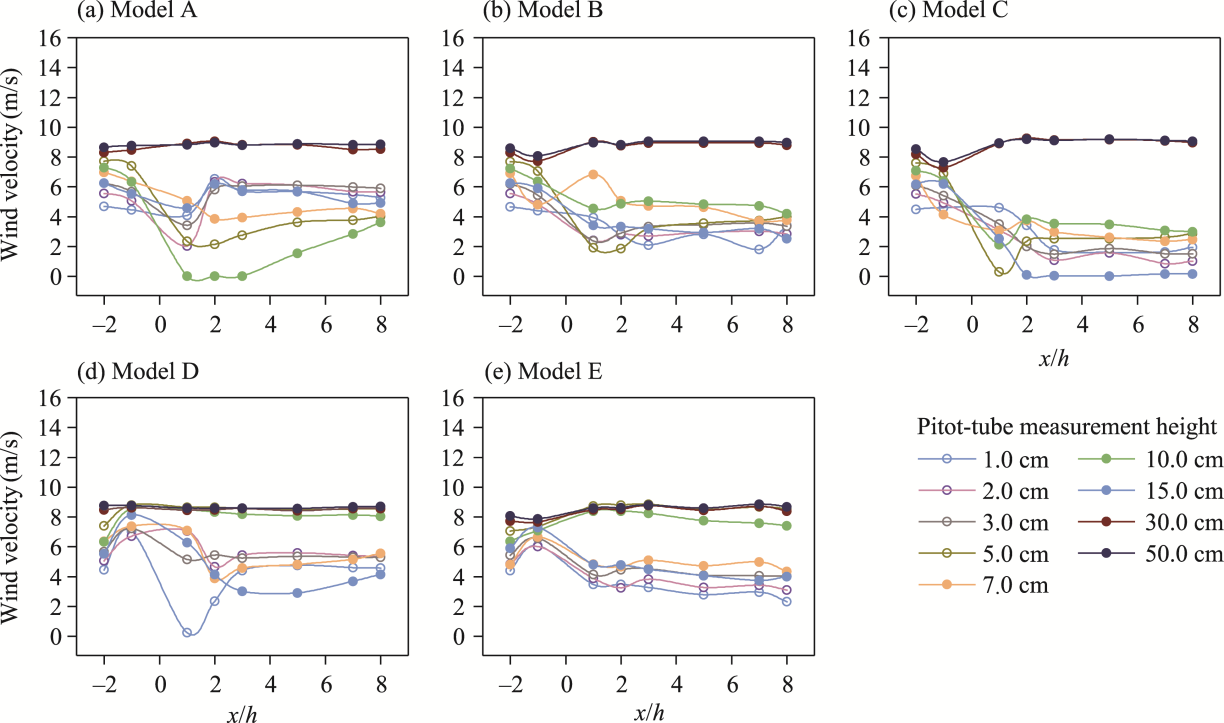
Fig. 5 Vertical wind velocity profiles at upwind and downwind positions for the five shelterbelt models A-E (a-e) under an initial wind velocity of 8.0 m/s. x/h is the normalized downwind distance, where x refers to the distance from the front edge (upwind side) of the shelterbelt for upwind measurements, and the distance from the rear edge (downwind side) for downwind measurements, and h represents the canopy height. Dashed horizontal lines indicate the canopy tops, where 2.0, 10.0, and 20.0 cm represent the canopy heights of C. esculentus, T. chinensis, and P. euphratica, respectively. Solid horizontal lines indicate the canopy bases, with 1.0 cm corresponding to the canopy base of C. esculentus and T. chinensis, and 10.0 cm corresponding to the canopy base of P. euphratica. Within the solid-line range, 15.0 cm denotes the height of the nylon net. |
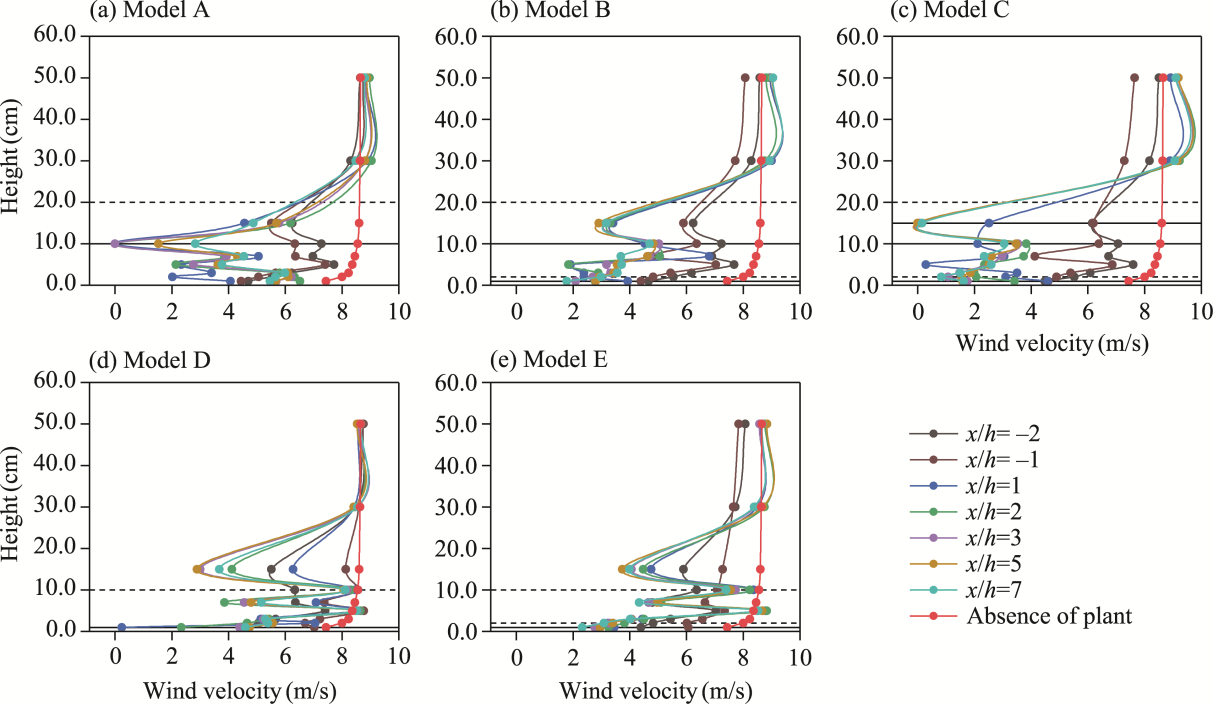
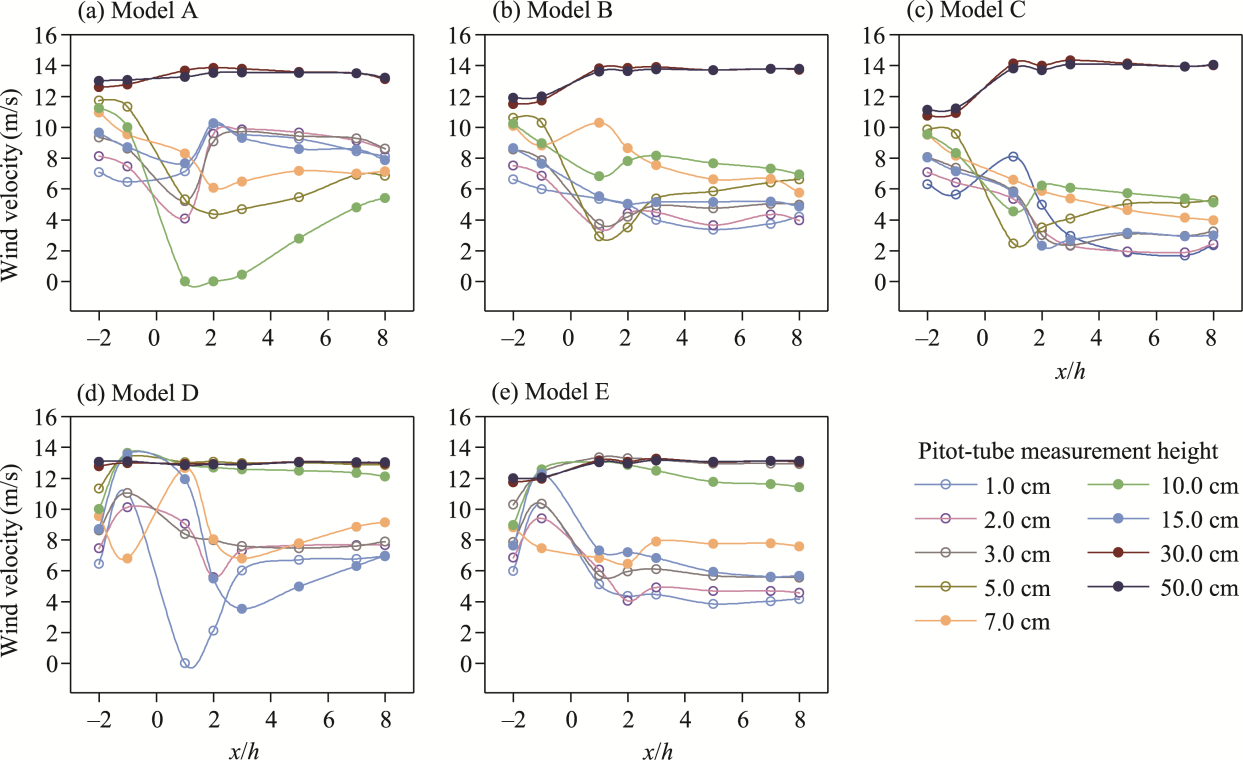
Fig. 6 Vertical wind velocity profiles at upwind and downwind positions for the five shelterbelt models A-E (a-e) under an initial wind velocity of 12.0 m/s. Dashed horizontal lines indicate the canopy tops, where 2.0, 10.0, and 20.0 cm represent the canopy heights of C. esculentus, T. chinensis, and P. euphratica, respectively. Solid horizontal lines indicate the canopy bases, with 1.0 cm corresponding to the canopy base of C. esculentus and T. chinensis, and 10.0 cm corresponding to the canopy base of P. euphratica. Within the solid-line range, 15.0 cm denotes the height of the nylon net. |
Fig. 7 Horizontal wind velocity profiles at upwind and downwind positions for five shelterbelt models A-E (a-e) under an initial wind velocity of 8.0 m/s |
Fig. 8 Horizontal wind velocity profiles at upwind and downwind positions for five shelterbelt models A-E (a-e) under an initial wind velocity of 12.0 m/s |
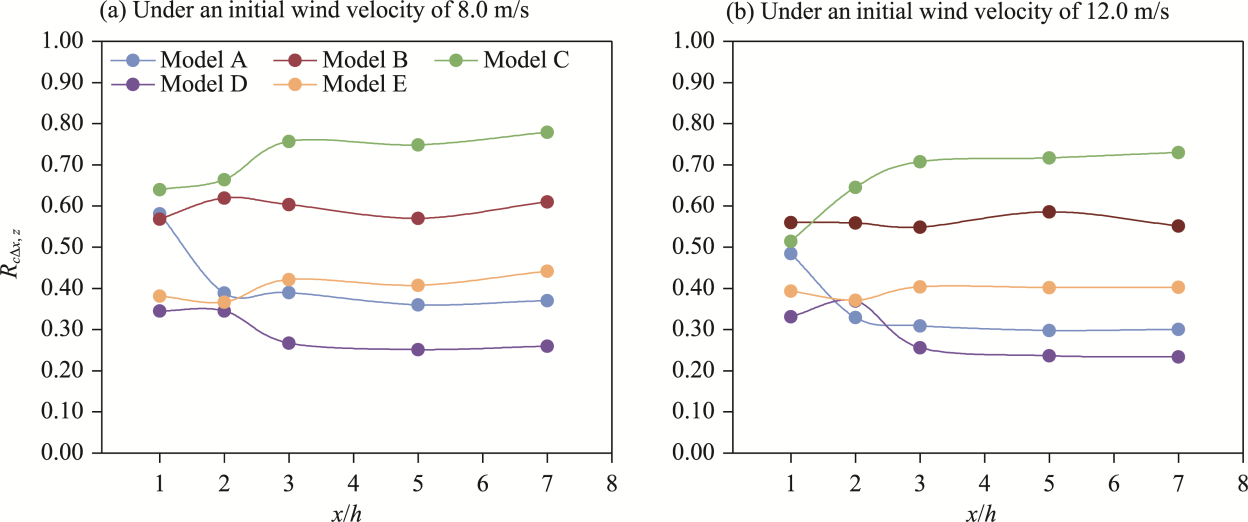
Fig. 9 Comparison of wind abatement coefficient (RcΔx,z) at different downwind distances among five shelterbelt models A-E under initial wind velocities of 8.0 m/s (a) and 12.0 m/s (b) |
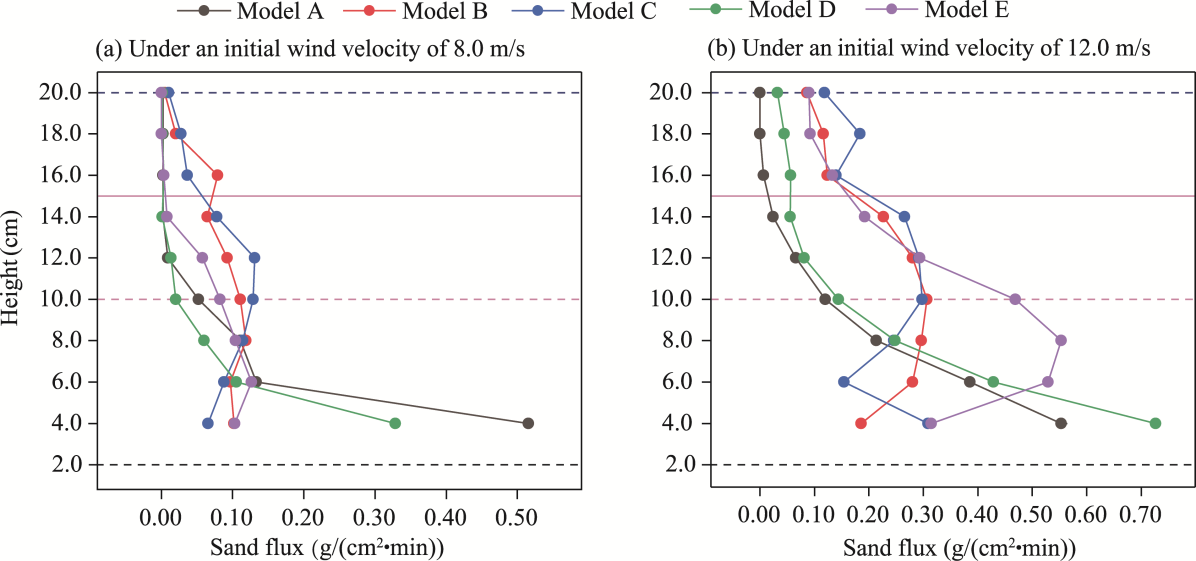
Fig. 10 Sand flux profiles for five shelterbelt models A-E under initial wind velocities of 8.0 m/s (a) and 12.0 m/s (b). Dashed horizontal lines indicate the canopy tops of the shelterbelt components, where 2.0, 10.0, and 20.0 cm represent the canopy heights of C. esculentus, T. chinensis, and P. euphratica, respectively. Within the range indicated by the solid horizontal lines, 15.0 cm denotes the height of the nylon net. |
Fig. 11 Comparison of cumulative sand flux within a layer extending from the ground surface to a height of 20.0 cm for five shelterbelt models A-E under initial wind velocities of 8.0 and 12.0 m/s. Bars are standard errors. |
Fig. 12 Variation in wind velocity at heights of 50.0, 100.0, and 200.0 cm in the C. esculentus plot compared to the bare sand plot (a) and comparison of cumulative sand flux at corresponding heights from July to September in the same experimental plots (b) |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
/
| 〈 |
|
〉 |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}