
Assessment of plant diversity in the Surkhan-Sherabad Region, Uzbekistan by grid mapping
Received date: 2024-10-16
Revised date: 2025-02-20
Accepted date: 2025-03-05
Online published: 2025-08-13
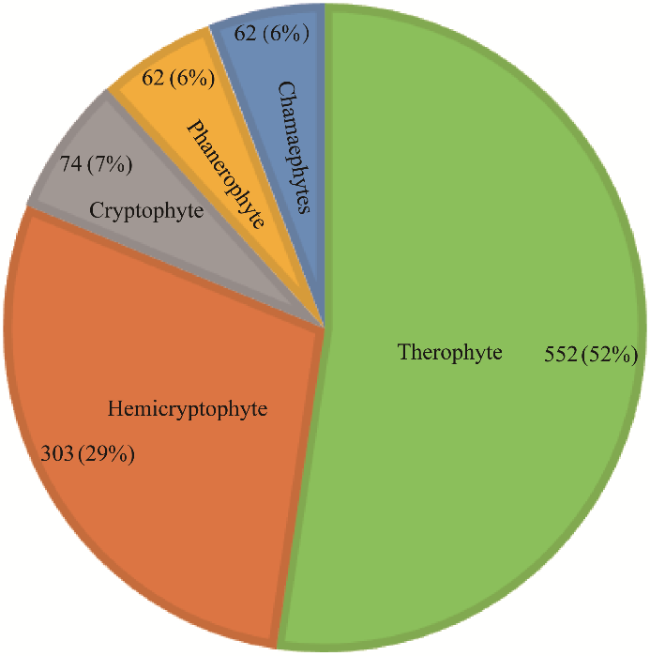
In floristic research, the grid mapping method is a crucial and highly effective tool for investigating the flora of specific regions. This methodology aids in the collection of comprehensive data, thereby promoting a thorough understanding of regional plant diversity. This paper presents findings from a grid mapping study conducted in the Surkhan-Sherabad botanical-geographic region (SShBGR), acknowledged as one of the major floristic areas in southwestern Uzbekistan. Using an expansive dataset of 14,317 records comprised of herbarium specimens and field diary entries collected from 1897 to 2023, we evaluated the stages and seasonal dynamics of data accumulation, species richness (SR), and collection density (CD) within 5 km×5 km grid cells. We further examined the taxonomic and life form composition of the region's flora. Our analysis revealed that the grid mapping phase (2021-2023) produced a significantly greater volume of specimens and taxonomic diversity compared with other periods (1897-1940, 1941-1993, and 1994-2020). Field research spanned 206 grid cells during 2021-2023, resulting in 11,883 samples, including 6469 herbarium specimens and 5414 field records. Overall, fieldwork covered 251 of the 253 grid cells within the SShBGR. Notably, the highest species diversity was documented in the B198 grid cell, recording 160 species. In terms of collection density, the E198 grid cell produced 475 samples. Overall, we identified 1053 species distributed across 439 genera and 78 families in the SShBGR. The flora of this region aligned significantly with the dominant families commonly found in the Holarctic, highlighting vital ecological connections. Among our findings, the Asteraceae family was the most polymorphic, with 147 species, followed by the continually stable and diverse Poaceae, Fabaceae, Brassicaceae, and Amaranthaceae. Besides, our analysis revealed a predominance of therophyte life forms, which constituted 52% (552 species) of the total flora. The findings underscore the necessity for continual data collection efforts to further enhance our understanding of the biodiversity in the SShBGR. The results of this study demonstrated that the application of grid-based mapping in floristic studies proves to be an effective tool for assessing biodiversity and identifying key taxonomic groups.
Inom JURAMURODOV , Rustam URALOV , Dilmurod MAKHMUDJANOV , LU Chunfang , Feruz AKBAROV , Sardor PULATOV , Bakhtiyor KARIMOV , Orzimat TURGINOV , Komiljon TOJIBAEV . Assessment of plant diversity in the Surkhan-Sherabad Region, Uzbekistan by grid mapping[J]. Journal of Arid Land, 2025 , 17(3) : 394 -410 . DOI: 10.1007/s40333-025-0096-5
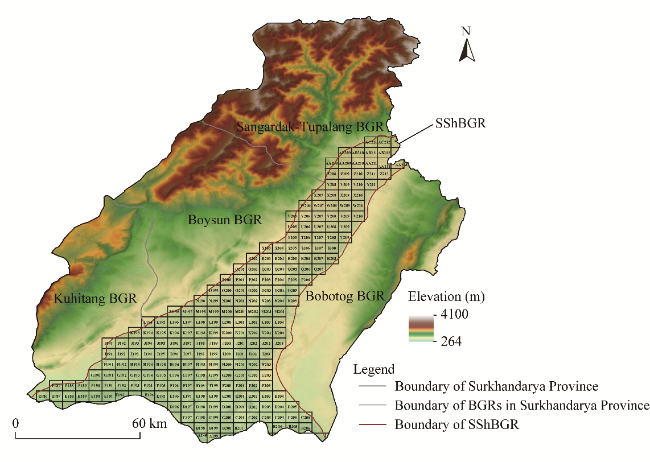
Fig. 1 Grid system established in the Surkhan-Sherabad botanical-geographic region (SShBGR). BGR, botanical-geographic region. |
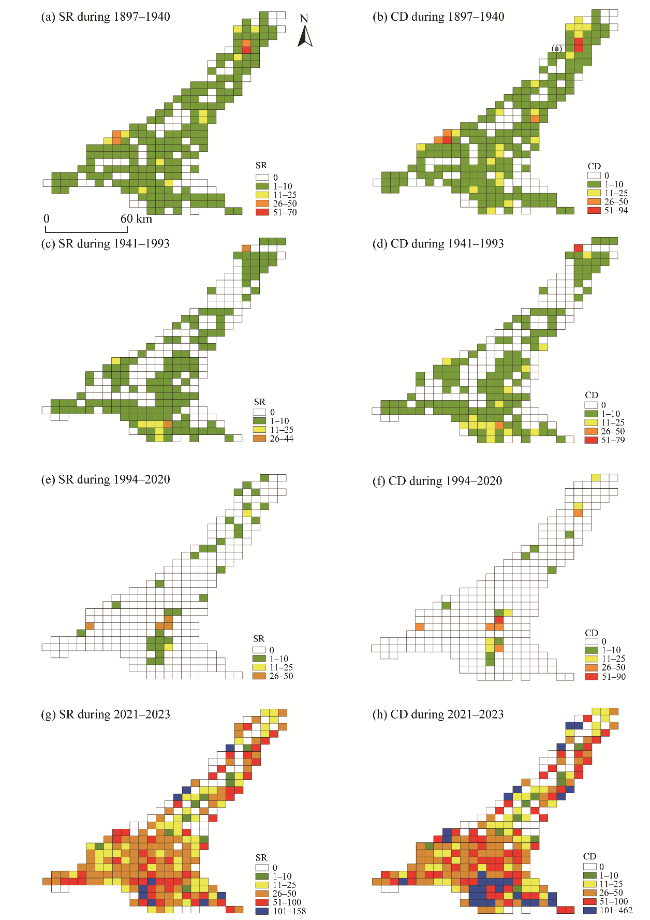
Fig. 2 Distribution of species richness (SR) and collection density (CD) for data collected from the grid system in the SShBGR across different periods. (a), SR during 1897-1940; (b), CD during 1897-1940; (c), SR during 1941-1993; (d), CD during 1941-1993; (e), SR during 1994-2020; (f), CD during 1994-2020; (g), SR during 2021-2023; (h), CD during 2021-2023. |
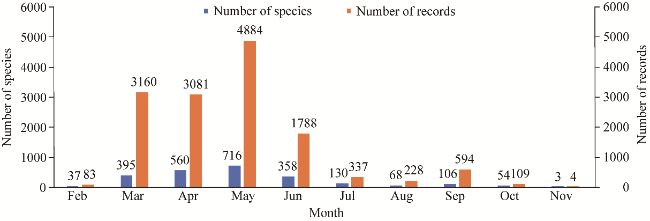
Fig. 3 Monthly dynamics of plant data collection from the SShBGR from 1897 to 2023 |
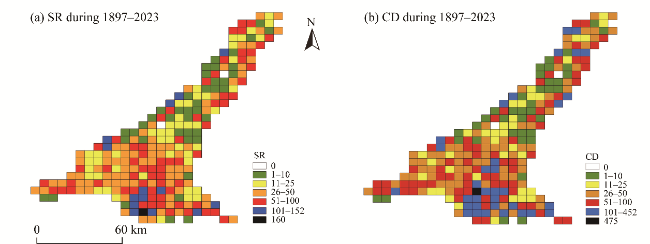
Fig. 4 Visualization of all species occurrence records collected from the SShBGR from 1897 to 2023. (a), SR during 1897-2023; (b), CD during 1897-2023. |
Table 1 Number of plant families, genera, species, and records collected from the SShBGR across different periods |
| Period | Number of families | Number of genera | Number of species | Number of records |
|---|---|---|---|---|
| Period A (1897-1940) | 54 | 244 | 428 | 1265 |
| Period B (1941-1993) | 49 | 181 | 299 | 800 |
| Period C (1994-2020) | 38 | 105 | 139 | 362 |
| Period D (2021-2023) | 65 | 367 | 803 | 11,883 |
Table 2 List of top-ten polymorphic families in the Surkhan-Sherabad botanical-geographic region (SShBGR) in each period |
| Period | Rank | Family | Number of genera | Number of species | Number of records |
|---|---|---|---|---|---|
| Period A (1897-1940) | 1 | Poaceae | 35 | 48 | 190 |
| 2 | Asteraceae | 25 | 45 | 137 | |
| 3 | Fabaceae | 13 | 42 | 148 | |
| 4 | Brassicaceae | 22 | 34 | 68 | |
| 5 | Amaranthaceae | 20 | 31 | 107 | |
| 6 | Lamiaceae | 13 | 18 | 37 | |
| 7 | Apiaceae | 11 | 18 | 52 | |
| 8 | Boraginaceae | 11 | 18 | 33 | |
| 9 | Cyperaceae | 7 | 18 | 44 | |
| 10 | Plantaginaceae | 6 | 12 | 31 | |
| Period B (1941-1993) | 1 | Poaceae | 28 | 40 | 114 |
| 2 | Asteraceae | 18 | 33 | 77 | |
| 3 | Fabaceae | 12 | 32 | 79 | |
| 4 | Amaranthaceae | 16 | 24 | 98 | |
| 5 | Cyperaceae | 5 | 13 | 42 | |
| 6 | Lamiaceae | 10 | 12 | 28 | |
| 7 | Brassicaceae | 10 | 11 | 14 | |
| 8 | Apiaceae | 9 | 11 | 20 | |
| 9 | Plantaginaceae | 3 | 7 | 10 | |
| 10 | Boraginaceae | 6 | 6 | 10 | |
| Period C (1994-2020) | 1 | Asteraceae | 15 | 18 | 38 |
| 2 | Brassicaceae | 11 | 12 | 46 | |
| 3 | Poaceae | 9 | 10 | 26 | |
| 4 | Fabaceae | 5 | 10 | 29 | |
| 5 | Liliaceae | 3 | 10 | 21 | |
| 6 | Amaranthaceae | 8 | 9 | 31 | |
| 7 | Lamiaceae | 8 | 8 | 13 | |
| 8 | Caryophyllaceae | 5 | 7 | 16 | |
| 9 | Ranunculaceae | 3 | 5 | 14 | |
| 10 | Plantaginaceae | 2 | 3 | 5 | |
| Period D (2021-2023) | 1 | Asteraceae | 50 | 122 | 1567 |
| 2 | Poaceae | 48 | 93 | 1847 | |
| 3 | Fabaceae | 19 | 73 | 1049 | |
| 4 | Amaranthaceae | 32 | 71 | 1025 | |
| 5 | Brassicaceae | 40 | 68 | 1432 | |
| 6 | Caryophyllaceae | 16 | 32 | 210 | |
| 7 | Boraginaceae | 14 | 29 | 396 | |
| 8 | Apiaceae | 23 | 28 | 181 | |
| 9 | Polygonaceae | 6 | 26 | 492 | |
| 10 | Ranunculaceae | 8 | 25 | 112 |
Table 3 Taxonomic and life forms of 14,317 records collected from the SShBGR from 1897 to 2023 |
| No. | Family | Number of genera | Number of species | Number of life forms | Number of records | ||||
|---|---|---|---|---|---|---|---|---|---|
| T | H | Cr | Ch | P | |||||
| 1 | Asteraceae | 58 | 147 | 85 | 48 | 1 | 13 | - | 1819 |
| 2 | Poaceae | 53 | 109 | 65 | 43 | 1 | - | - | 2178 |
| 3 | Fabaceae | 20 | 96 | 44 | 38 | - | 11 | 3 | 1305 |
| 4 | Brassicaceae | 45 | 80 | 73 | 7 | - | - | - | 1561 |
| 5 | Amaranthaceae | 33 | 79 | 59 | - | - | 13 | 7 | 1261 |
| 6 | Lamiaceae | 20 | 42 | 10 | 21 | - | 10 | 1 | 352 |
| 7 | Caryophyllaceae | 17 | 40 | 29 | 11 | - | - | - | 251 |
| 8 | Apiaceae | 29 | 39 | 16 | 11 | 12 | - | - | 255 |
| 9 | Boraginaceae | 16 | 39 | 27 | 12 | - | - | - | 439 |
| 10 | Polygonaceae | 7 | 28 | 9 | 6 | - | - | 13 | 567 |
| 11 | Ranunculaceae | 8 | 28 | 15 | 12 | - | - | 1 | 158 |
| 12 | Cyperaceae | 9 | 26 | 9 | 17 | - | - | - | 306 |
| 13 | Plantaginaceae | 7 | 25 | 14 | 11 | - | - | - | 502 |
| 14 | Liliaceae | 3 | 18 | - | - | 18 | - | - | 67 |
| 15 | Euphorbiaceae | 2 | 14 | 13 | 1 | - | - | - | 127 |
| 16 | Rubiaceae | 6 | 14 | 9 | 3 | - | 2 | - | 76 |
| 17 | Convolvulaceae | 4 | 13 | 3 | 7 | - | 3 | - | 340 |
| 18 | Tamaricaceae | 3 | 13 | - | - | - | 2 | 11 | 333 |
| 19 | Amaryllidaceae | 1 | 12 | - | - | 12 | - | - | 59 |
| 20 | Caprifoliaceae | 4 | 12 | 10 | 2 | - | - | - | 64 |
| 21 | Malvaceae | 5 | 12 | 6 | 6 | - | - | - | 225 |
| 22 | Rosaceae | 7 | 10 | - | 5 | - | - | 5 | 31 |
| 23 | Papaveraceae | 5 | 9 | 9 | - | - | - | - | 295 |
| 24 | Geraniaceae | 2 | 9 | 8 | 1 | - | - | - | 266 |
| 25 | Solanaceae | 4 | 9 | 5 | - | - | 1 | 3 | 113 |
| 26 | Zygophyllaceae | 2 | 7 | 2 | 2 | - | 3 | - | 233 |
| 27 | Iridaceae | 1 | 6 | - | - | 6 | - | - | 55 |
| 28 | Juncaceae | 1 | 6 | - | 6 | - | - | - | 15 |
| 29 | Rutaceae | 1 | 6 | - | 6 | - | - | - | 74 |
| 30 | Typhaceae | 1 | 6 | - | - | 6 | - | - | 74 |
| 31 | Asparagaceae | 4 | 5 | - | - | 5 | - | - | 12 |
| 32 | Ephedraceae | 1 | 5 | - | - | - | - | 5 | 38 |
| 33 | Lythraceae | 3 | 5 | 4 | 1 | - | - | - | 22 |
| 34 | Orobanchaceae | 2 | 5 | - | 5 | - | - | - | 19 |
| 35 | Salicaceae | 2 | 5 | - | - | - | - | 5 | 39 |
| 36 | Scrophulariaceae | 2 | 5 | 2 | 3 | - | - | - | 39 |
| 37 | Plumbaginaceae | 2 | 3 | 2 | 1 | - | - | - | 32 |
| 38 | Apocynaceae | 2 | 3 | - | 3 | - | - | - | 71 |
| 39 | Araceae | 2 | 3 | - | - | 3 | - | - | 41 |
| 40 | Asphodelaceae | 1 | 3 | - | - | 3 | - | - | 4 |
| 41 | Cleomaceae | 1 | 3 | 3 | - | - | - | - | 6 |
| 42 | Frankeniaceae | 1 | 3 | 1 | - | - | 2 | - | 10 |
| 43 | Onagraceae | 2 | 3 | 1 | 2 | - | - | - | 6 |
| 44 | Orchidaceae | 3 | 3 | - | - | 3 | - | - | 6 |
| 45 | Primulaceae | 1 | 3 | 3 | - | - | - | - | 34 |
| 46 | Crassulaceae | 1 | 1 | 1 | - | - | - | - | 1 |
| 47 | Cynomoriaceae | 1 | 2 | 2 | - | - | - | - | 5 |
| 48 | Equisetaceae | 1 | 2 | - | - | 2 | - | - | 34 |
| 49 | Gentianaceae | 2 | 2 | 1 | 1 | - | - | - | 22 |
| 50 | Moraceae | 1 | 2 | - | - | - | - | 2 | 23 |
| 51 | Nitrariaceae | 2 | 2 | 1 | 1 | - | - | - | 90 |
| 52 | Phyllanthaceae | 1 | 2 | - | - | - | 2 | - | 4 |
| 53 | Potamogetonaceae | 1 | 2 | - | 2 | - | - | - | 4 |
| 54 | Resedaceae | 1 | 2 | 2 | - | - | - | - | 5 |
| 55 | Thymelaeaceae | 2 | 2 | 2 | - | - | - | - | 15 |
| 56 | Anacardiaceae | 1 | 1 | - | - | - | - | 1 | 3 |
| 57 | Balsaminaceae | 1 | 1 | 1 | - | - | - | - | 4 |
| 58 | Berberidaceae | 1 | 1 | - | 1 | - | - | - | 2 |
| 59 | Capparaceae | 1 | 1 | - | 1 | - | - | - | 47 |
| 60 | Colchicaceae | 1 | 1 | - | - | 1 | - | - | 47 |
| 61 | Datiscaceae | 1 | 1 | - | 1 | - | - | - | 1 |
| 62 | Elaeagnaceae | 1 | 1 | - | - | - | - | 1 | 38 |
| 63 | Elatinaceae | 1 | 1 | 1 | - | - | - | - | 2 |
| 64 | Eriocaulaceae | 1 | 1 | 1 | - | - | - | - | 1 |
| 65 | Hypericaceae | 1 | 1 | - | 1 | - | - | - | 4 |
| 66 | Ixioliriaceae | 1 | 1 | - | - | 1 | - | - | 50 |
| 67 | Linaceae | 1 | 1 | - | 1 | - | - | - | 4 |
| 68 | Mazaceae | 1 | 1 | - | 1 | - | - | - | 30 |
| 69 | Oleaceae | 1 | 1 | - | - | - | - | 1 | 1 |
| 70 | Oxalidaceae | 1 | 1 | 1 | - | - | - | - | 20 |
| 71 | Portulacaceae | 1 | 1 | 1 | - | - | - | - | 53 |
| 72 | Pteridaceae | 1 | 1 | - | 1 | - | - | - | 1 |
| 73 | Sapindaceae | 1 | 1 | - | - | - | - | 1 | 2 |
| 74 | Sphenocleaceae | 1 | 1 | 1 | - | - | - | - | 5 |
| 75 | Ulmaceae | 1 | 1 | - | - | - | - | 1 | 7 |
| 76 | Urticaceae | 1 | 1 | 1 | - | - | - | - | 1 |
| 77 | Verbenaceae | 1 | 1 | - | 1 | - | - | - | 10 |
| 78 | Vitaceae | 1 | 1 | 1 | - | - | - | 1 | 1 |
| Total | 439 | 1053 | 552 | 303 | 74 | 62 | 62 | 14,317 | |
Note: T, therophyte; H, hemicryptophyte; Cr, cryptophyte; Ch, chamaephyte; P, phanerophyte. ''-'' indicates no value. |
Fig. 5 Number and proportion of life form of the flora in the SShBGR |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}