
Extreme drought with seasonal timing consistently promotes CH4 uptake through inconsistent pathways in a temperate grassland, China
Received date: 2024-03-05
Revised date: 2024-04-23
Accepted date: 2024-05-15
Online published: 2025-08-13
Methane (CH4) is a potent greenhouse gas that has a substantial impact on global warming due to its substantial influence on the greenhouse effect. Increasing extreme precipitation events, such as drought, attributable to global warming that caused by greenhouse gases, exert a profound impact on the intricate biological processes associated with CH4 uptake. Notably, the timing of extreme drought occurrence emerges as a pivotal factor influencing CH4 uptake, even when the degree of drought remains constant. However, it is still unclear how the growing season regulates the response of CH4 uptake to extreme drought. In an effort to bridge this knowledge gap, we conducted a field manipulative experiment to evaluate the impact of extreme drought on CH4 uptake during early, middle, and late growing stages in a temperate steppe of Inner Mongolia Autonomous Region, China. The result showed that all extreme drought consistently exerted positive effects on CH4 uptake regardless of seasonal timing. However, the magnitude of this effect varied depending on the timing of season, as evidenced by a stronger effect in early growing stage than in middle and late growing stages. Besides, the pathways of CH4 uptake were different from seasonal timing. Extreme drought affected soil physical-chemical properties and aboveground biomass (AGB), consequently leading to changes in CH4 uptake. The structural equation model showed that drought both in the early and middle growing stages enhanced CH4 uptake due to reduced soil water content (SWC), leading to a decrease in NO3--N and an increase in pmoA abundance. However, drought in late growing stage primarily enhanced CH4 uptake only by decreasing SWC. Our results suggested that seasonal timing significantly contributed to regulate the impacts of extreme drought pathways and magnitudes on CH4 uptake. The findings can provide substantial implications for understanding how extreme droughts affect CH4 uptake and improve the prediction of potential ecological consequence under future climate change.
Key words: extreme climate; greenhouse gas; methane; methanotrophs; soil inorganic nitrogen
ZHANG Wenwen , PAN Yue , WEN Fuqi , FU Juanjuan , HAO Yanbin , HU Tianming , YANG Peizhi . Extreme drought with seasonal timing consistently promotes CH4 uptake through inconsistent pathways in a temperate grassland, China[J]. Journal of Arid Land, 2024 , 16(6) : 768 -778 . DOI: 10.1007/s40333-024-0017-z
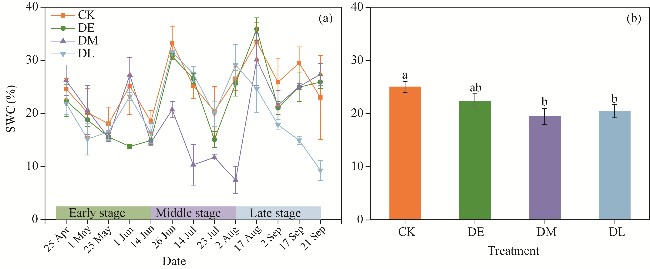
Fig. 1 Variations in seasonal (a) and average (b) soil water content (SWC) under different drought treatments. Different lowercase letters in Figure 1b indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. DE, DM, and DL are droughts in early, middle, and late growing stages, respectively. CK, control. The abbreviations are the same in the following figures. |
Table 1 Effects of drought in early, middle, and late growing stages on SWC, NH4+-N, NO3--N, AGB, CH4 uptake, and pmoA abundance |
| Treatment | SWC | NH4+-N | NO3--N | |||
|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |
| DE | 4.827 | 0.0370 | 8.272 | 0.0210 | 3.195 | 0.0410 |
| DM | 5.686 | 0.0250 | 0.113 | 0.7450 | 1.698 | 0.0300 |
| DL | 2.844 | 0.0700 | 1.471 | 0.2590 | 1.094 | 0.3260 |
| Treatment | AGB | CH4 uptake | pmoA | |||
| F | P | F | P | F | P | |
| DE | 0.055 | 0.0550 | 6.292 | 0.0370 | 28.103 | 0.0410 |
| DM | 1.338 | 0.2820 | 7.455 | 0.0480 | 87.063 | <0.0001 |
| DL | 7.345 | 0.0270 | 15.258 | 0.0400 | 45.627 | 0.0001 |
Note: DE, DM, and DL are droughts in early, middle, and late growing stages, respectively; SWC, soil water content; AGB, aboveground biomass; CH4, methane. |
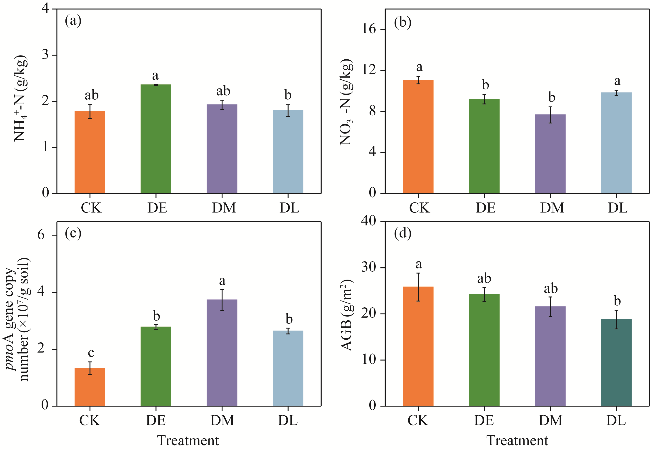
Fig. 2 Variations in NH4+-N (a), NO3--N (b), pmoA gene copy number (c), and AGB (d) under different drought treatments. AGB, aboveground biomass. Different lowercase letters indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. |
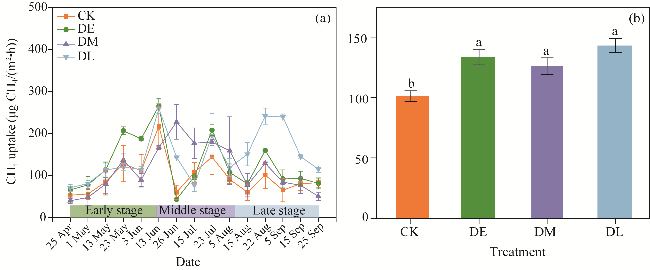
Fig. 3 Seasonal dynamics (a) and mean (b) of CH4 uptake under different drought treatments. Different lowercase letters in Figure 3b indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. |
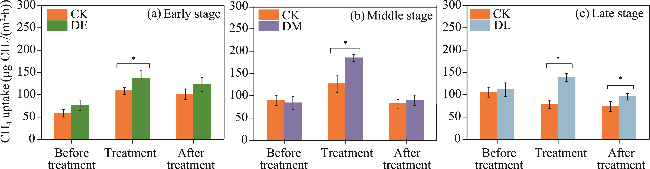
Fig. 4 CH4 uptake before, during, and after drought treatments. (a), drought in early growing stage; (b), drought in middle growing stage; (c), drought in late growing stage. * indicates significant differences between treatments at P<0.05 level. Bars are standard errors. |
Table 2 Multiple regression analyses of drought on CH4 uptake and pmoA abundance |
| Regression equation | R2 | P | SEDE | SEDM |
|---|---|---|---|---|
| CH4 uptake=124.80+1.15SWC-0.98AGB+17.23DE+13.86DM | 0.973 | 0.02 | 7.28 | 2.99 |
| pmoA=3.25×107-1.72×106SWC+9.01×105AGB-1.82×107DE+3.27×107DM | 0.989 | 0.02 | 1.42×107 | 1.10×107 |
Note: SE, standardized regression coefficient; DE and DM are droughts in early and middle growing stages, respectively. |
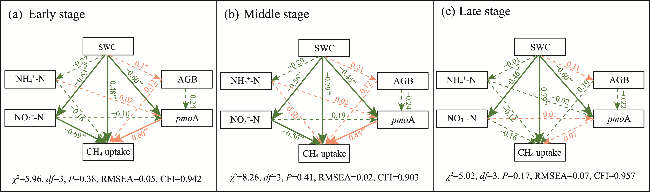
Fig. 5 Structural equation model (SEM) showing effects of abiotic factors and biotic factors on CH4 uptake under drought in early (a), middle (b), and late (c) growing stages. Line with orange color indicates positive path coefficients, and line with green color indicates negative path coefficients. Solid and dashed lines represent significant and insignificant path coefficients, respectively. RMSEA, root mean square error of approximation; CFI, comparative fit index. *, significant at P<0.05 level; **, significant at P<0.01 level. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
IPCC (Intergovernmental Panel on Climate Change). 2013. The physical science basis. In: Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
|
| [26] |
IPCC (Intergovernmental Panel on Climate Change). 2021. Climate change 2021:The physical science basis. In: Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}