
Diversification of flavonoid accumulation among ecotypes of Agriophyllum squarrosum (L.) Moq. in response to drought stress
Received date: 2024-08-26
Revised date: 2025-01-25
Accepted date: 2025-02-26
Online published: 2025-08-13
Agriophyllum squarrosum (L.) Moq., commonly known as sandrice, is an annual medicinal plant prevalent in the dunes across China's deserts. A garden trial revealed that flavonoid content varies among sandrice ecotypes due to long-term local adaptation to water variability. To investigate how sandrice responds to drought stress through the molecular metabolic regulation of flavonoids, we employed transcriptomic and metabolomic analyses during a 9-d ambient drought stress, examining three ecotypes along a precipitation gradient. The three ecotypes located in Dengkou (DK) County, Dulan (DL) County, and Aerxiang (AEX) village of northern China, which had 137, 263, and 485 mm precipitation, respectively. Soil moisture content was 4.04% after drought stress, causing seedlings of the three sandrice ecotypes to display collapsed structures, yellowing leaves, wilting, and curling. Among these, DL exhibited superior drought tolerance, in which plant height increase (PHI) and leaf area (LA) were significantly higher than those of DK and AEX. Flavonoid-targeted metabolomics identified that rutin, isoquercitrin, and astragalin constituted over 95.00% of the 15 flavonoid metabolites detected. A total of 12 differentially accumulated flavonoids (DAFs) were found, with rutin being the most abundant (1231.57-2859.34 ng/100 mg fresh weight (FW)), showing a gradual increase along the precipitation gradient. Transcriptomic analysis revealed 14 common differentially expressed genes (DEGs) associated with flavonoid synthesis among the three ecotypes. Integrative analysis of DEGs and DAFs indicated that sandrice adapts to drought stress by activating different flavonoid synthesis pathways. In DK, the dihydrokaempferol-dihydroquercetin pathway, regulated by flavonoid 3'-monooxygenase (CYP75B1), likely enhances drought adaptation. In AEX, transcriptional repression by O-methylatransferase (OMT) shifts the metabolic flux from the quercetin-isorhamnetin pathway to the quercetin-isoquercetin-rutin pathway in response to drought. DL, the most drought- tolerant ecotype, appears to activate the naringenin-apigenin-luteolin route and employs a unique flavonoid accumulation pattern in response to drought stress. Our data reveal that flavonoid synthesis in sandrice is fine-tuned among ecotypes to cope with drought, offering valuable germplasm resources and evaluation methods for sandrice acclimation and providing insights into drought response in non-model plants.
Key words: drought tolerance; ecotype; flavonoid; medicinal plant; rutin
ZHAO Pengshu , YAN Xia , QIAN Chaoju , MA Guorong , FANG Tingzhou , YIN Xiaoyue , ZHOU Shanshan , LIAO Yuqiu , SHI Liang , FAN Xingke , Awuku IBRAHIM , MA Xiaofei . Diversification of flavonoid accumulation among ecotypes of Agriophyllum squarrosum (L.) Moq. in response to drought stress[J]. Journal of Arid Land, 2025 , 17(4) : 538 -559 . DOI: 10.1007/s40333-025-0011-0
Table 1 Native habitat environments and phenotypic characteristics of the three ecotypes |
| Ecotype | Collection site | Altitude (m) | Latitude | Longitude | Mean annual precipitation (mm) | Annual mean temperature (°C) | Phenotypic characteristics |
|---|---|---|---|---|---|---|---|
| DK | Dengkou | 1050 | 40°22°42"N | 106°59°37"E | 137 | 8.31 | Small seed size and short broad leaf |
| DL | Dulan | 3130 | 36°25°25"N | 98°07°25"E | 263 | 3.40 | Seed wing, long broad thick leaf, and large leaf area |
| AEX | Aerxiang | 251 | 42°52°04"N | 122°25°40"E | 485 | 6.38 | Large seed size and long thin leaf |
Note: DK, Dengkou County; DL, Dulan County; AEX, Aerxiang Village. |
Table S1 Primers for quantitative real-time PCR (qRT-PCR) analysis |
| Gene name | Gene No. | Forward primer (5° to 3°) | Reverse primer (5° to 3°) | Primer melting temperature (°C) | Product length (bp) |
|---|---|---|---|---|---|
| UBC22 | AsqAEX016065 | AATGGAGCCCAGCACTTACA | GCCTCATACTTTGGCCGATCA | 57.00 | 133 |
| PP2A | AsqAEX007496 | TCGCCCCTGTTTAGTGGAGT | GTACGTCTTCTCTGCCGACT | 58.00 | 122 |
| 3'GT | AsqAEX018709 | TAGCAAAGCTCTGGGCATCC | GTGAGTCTATGGAGTCGCGG | 58.00 | 90 |
| CYP75B1 | AsqAEX001259 | CGGAGGTGCTAAAGAAGGCA | GACTACAGGGTGCAAACGGA | 58.00 | 137 |
| ANR | AsqAEX017402 | ACTCCCAGCCCTAACTGCTA | CTCCACCCCAGCTACTGTTG | 58.00 | 132 |
| CHS | AsqAEX006531 | GGGGTCAGCCCAAGTCTAAG | TTGGTGAGTTGGTAGTCCGC | 58.00 | 88 |
| FLS | AsqAEX017561 | GACTGGGGCTGCAAGAAGAT | ATCAGGGCAAGGACATGGTG | 58.00 | 101 |
| COMT5 | AsqAEX006593 | CTACAGAATCCGGATGCCCC | CGACTGTGCGAACTGTTGTG | 58.00 | 94 |
| COMT1 | AsqAEX003966 | GCACATTTGCCAGCCAAGAA | GGTCCTGATGGTGCAAGTGA | 58.00 | 99 |
| CHI | AsqAEX010035 | TCTGGAAGGCCATCGGAGTA | AAAATAGAGTGGCCCGGTGG | 58.00 | 103 |
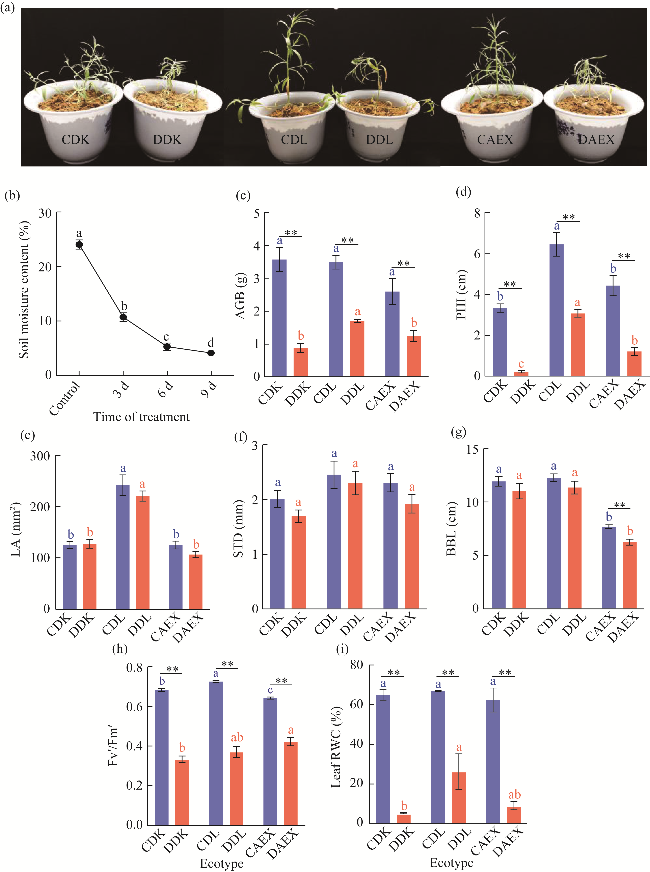
Fig. 1 Effects of drought stress on the morphological traits and physiological indicators in DK (Dengkou County), DL (Dulan County), and AEX (Aerxiang Village) ecotypes. (a), phenotype of DK, DL, and AEX after drought stress and control treatments (CDK, CDL, and CAEX); (b), soil moisture content; (c), above-ground biomass (AGB); (d), plant height increase (PHI); (e), leaf area (LA); (f), stem diameter (STD); (g), basal branch length (BBL); (h), Fv'/Fm' value at vegetative stage; (i), leaf relative water content (RWC). DDK, DDL, and DAEX represented the drought stress treatments of DK, DL and AEX, respectively. Bars are standard errors. Within the same treatment (control or drought), different lowercase letters indicated significant difference at P<0.050 level. Asterisks on shoulder lines indicated statistically significant differences between control and drought of the same ecotype. *, P<0.050 level; **, P<0.010 level. The abbreviations are the same in the following figures and tables. |
Table 2 Composition and average content of 15 flavonoids targeting metabolites |
| Compound name | CDK | DDK | CDL | DDL | CAEX | DAEX |
|---|---|---|---|---|---|---|
| (ng/100 mg fresh weight (FW)) | ||||||
| Rutin | 1231.57 | 1350.47 | 1668.45 | 1720.60 | 2495.48 | 2859.34 |
| Isoquercitrin | 1653.85 | 1442.55 | 1726.87 | 1461.72 | 1577.22 | 1593.38 |
| Astragalin | 818.28 | 714.61 | 741.53 | 519.81 | 650.12 | 722.03 |
| Quercetin | 12.48 | 14.44 | 10.68 | 7.91 | 17.55 | 26.85 |
| Vitexin | 12.06 | 26.72 | 14.93 | 21.38 | 4.28 | 10.27 |
| Isorhamnetin | 3.45 | 5.82 | 2.01 | 1.99 | 12.88 | 5.27 |
| Dihydrokaempferol | 1.16 | 1.93 | 2.12 | 5.13 | 2.78 | 3.49 |
| Kaempferol | 2.17 | 1.59 | 1.73 | 1.52 | 2.63 | 2.13 |
| Dihydroquercetin | 1.02 | 1.24 | 0.85 | 1.49 | 1.09 | 0.97 |
| Luteolin | 0.38 | 0.55 | 0.65 | 1.72 | 0.45 | 0.89 |
| Naringenin | 0.39 | 0.50 | 0.45 | 1.75 | 0.18 | 0.49 |
| Apigenin | 0.11 | 0.13 | 0.10 | 0.14 | 0.08 | 0.08 |
| Catechin | 0.41 | 0.13 | 0.07 | 0.11 | 0.16 | 0.07 |
| Epicatechin | 0.10 | 0.01 | 0.11 | 0.01 | 0.03 | 0.06 |
| Naringenin chalcone | 0.56 | 0.01 | 0.82 | 0.68 | 0.01 | 0.01 |
| Total flavonoid | 3737.99 | 3560.70 | 4171.37 | 3745.96 | 4764.94 | 5225.33 |
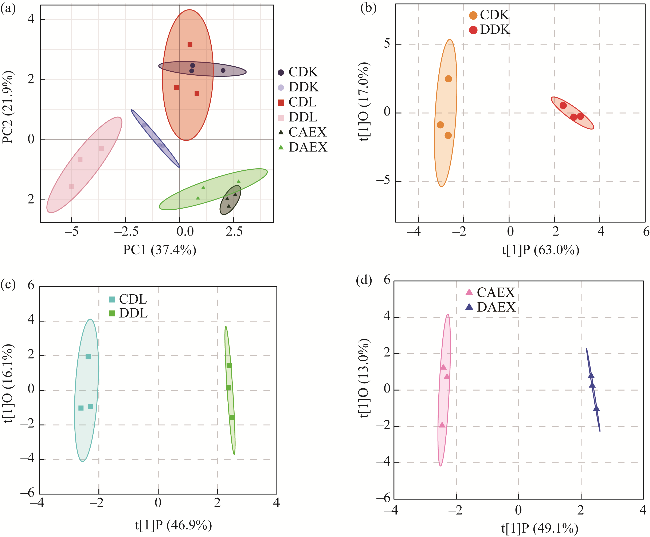
Fig. S1 Significant difference analysis of flavonoid-targeting metabolites in above-ground tissue of sandrice. (a), principal component analysis (PCA); (b-d), orthogonal partial least squares discriminant analysis (OPLS-DA). PC1 represents the first principal component, and PC2 represents the second principal component. t[1]P represents the predicted principal component score of the first principal component, showing the differences between sample groups, and t[1]O represents the orthogonal principal component score of the second principal component, showing the differences within sample groups. |
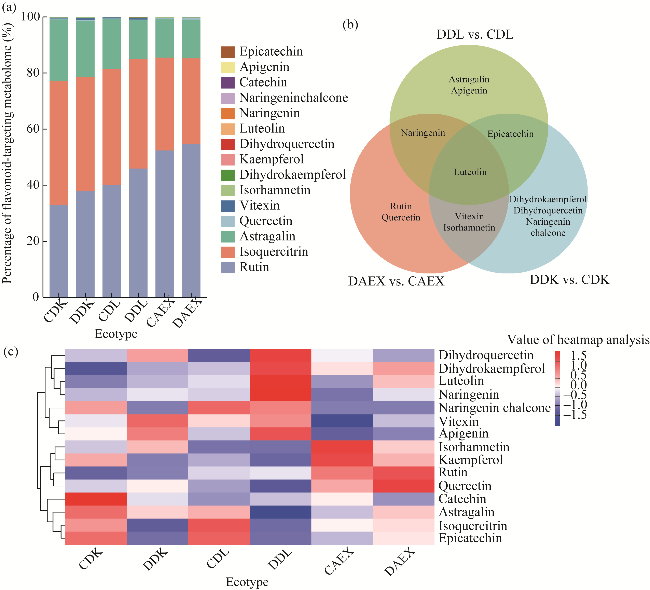
Fig. 2 Flavonoid-targeting metabolites identified in above-ground tissue of sandrice. (a), percentage of flavonoid-targeting metabolome; (b), Venn of metabolite significance analysis; (c), hierarchical cluster heatmap analysis of metabolites. |
Table S2 Summary of the ribonucleic acid sequence (RNA-Seq) results of sandrice aboveground tissue |
| Sample | Raw data (Gb) | Clean data (Gb) | Q30 (%) | Uniquely mapped (%) | Total mapped reads (%) | GC (%) |
|---|---|---|---|---|---|---|
| CDK1 | 7.09 | 6.68 | 91.05 | 93.16 | 96.25 | 44.78 |
| CDK2 | 7.21 | 6.71 | 91.20 | 93.53 | 96.38 | 45.21 |
| CDK3 | 7.61 | 7.08 | 90.87 | 93.21 | 96.11 | 45.09 |
| DDK1 | 6.83 | 6.38 | 90.69 | 92.19 | 95.72 | 44.37 |
| DDK2 | 6.91 | 6.47 | 90.55 | 92.23 | 95.68 | 44.25 |
| DDK3 | 6.93 | 6.45 | 90.77 | 92.48 | 95.78 | 44.43 |
| CDL1 | 7.01 | 6.63 | 91.91 | 93.40 | 96.22 | 45.28 |
| CDL2 | 6.66 | 6.22 | 91.92 | 93.45 | 96.17 | 45.12 |
| CDL3 | 7.41 | 6.95 | 91.04 | 93.06 | 95.89 | 44.68 |
| DDL1 | 7.32 | 6.91 | 91.32 | 92.52 | 95.71 | 44.47 |
| DDL2 | 7.63 | 7.19 | 91.21 | 92.57 | 95.87 | 44.37 |
| DDL3 | 7.29 | 6.88 | 91.57 | 92.68 | 96.16 | 44.38 |
| CAEX1 | 7.14 | 6.57 | 91.50 | 94.72 | 97.76 | 44.96 |
| CAEX2 | 7.11 | 6.70 | 91.32 | 94.56 | 97.55 | 44.94 |
| CAEX3 | 7.12 | 6.75 | 91.50 | 94.69 | 97.79 | 44.71 |
| DAEX1 | 7.34 | 6.87 | 91.22 | 94.39 | 97.78 | 44.37 |
| DAEX2 | 6.97 | 6.47 | 90.54 | 93.87 | 97.04 | 44.56 |
| DAEX3 | 6.97 | 6.53 | 90.78 | 94.34 | 97.62 | 44.49 |
Note: Q30 is the percentage of bases with a mass value greater than or equal to 30, and the higher the value, the better the quality of the sequencing. GC content refers to the total proportion of guanine (G) and cytosine (C) bases in a DNA sequence, usually expressed as a percentage. DK, Dengkou County; DL, Dulan County; AEX, Aerxiang Village. CDK, CDL, CAEX are the control treatments of DK, DL, and AEX, respectively; DDK, DDL, and DAEX are the drought stress treatments of DK, DL and AEX, respectively. |
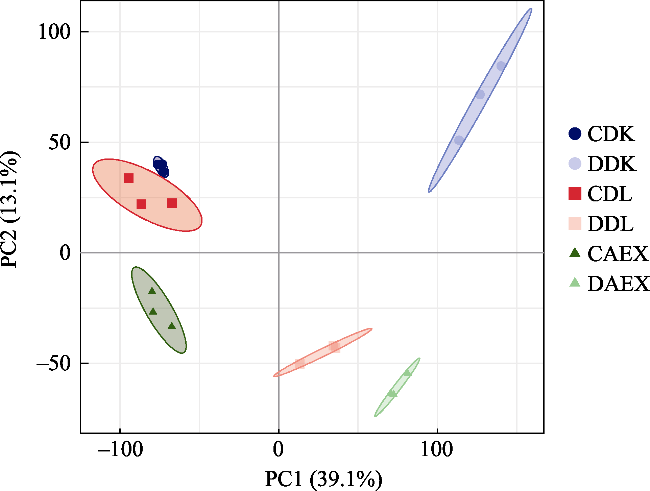
Fig. S2 PCA analysis showing the transcriptomes divergence |
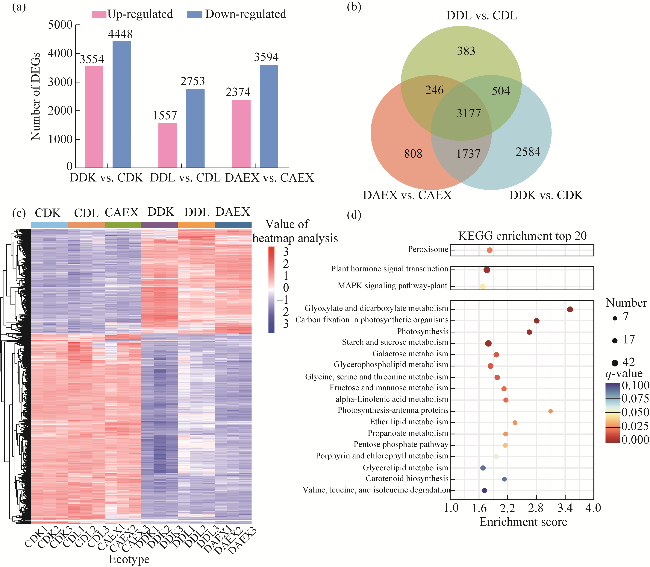
Fig. 3 Description of differentially expressed genes (DEGs) in transcriptome data of the three ecotypes of sandrice in response to drought. (a), number of DEGs; (b), Venn diagram showing overlap of DEGs in the three ecotypes; (c), heatmap analysis of 3177 DEGs in Figure 3b; (d), Kyoto encyclopedia of genes and genomes (KEGG) enrichment analysis bubble map. Dot size represents the number of distinct genes, and dot color reflects the q-value. |
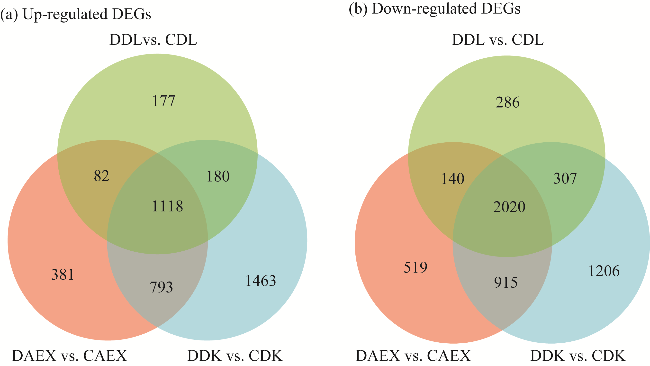
Fig. S3 Venn diagram showing overlap of up-regulated differentially expressed genes (DEGs) (a) and down-regulated DEGs (b) in the three ecotypes |
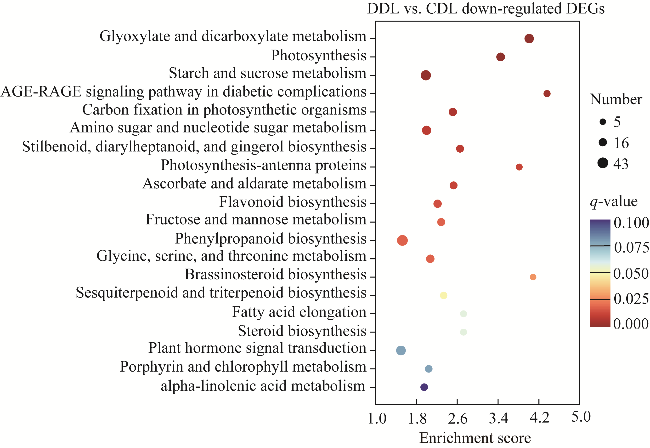
Fig. S4 Kyoto encyclopedia of genes and genomes (KEGG) enrichment analysis bubble map in DL. AGE-RAGE, advanced glycation end products-receptor for advanced glycation end products. |
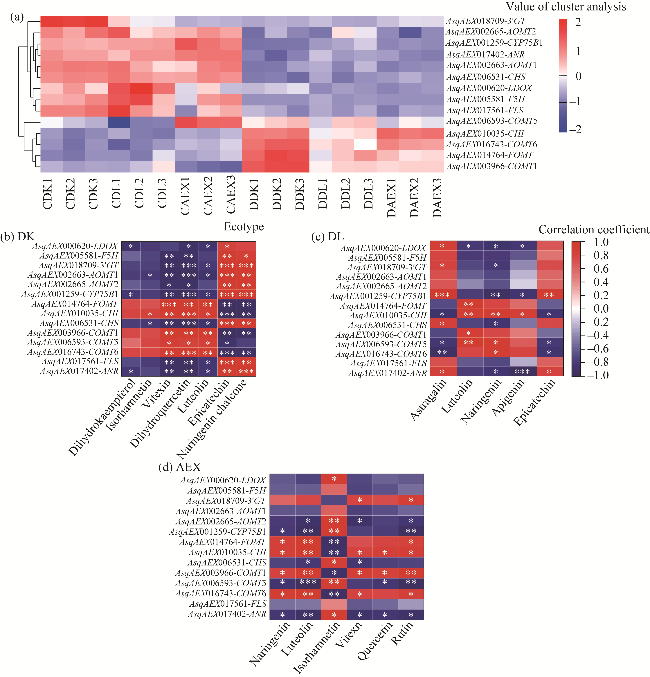
Fig. 4 Cluster analysis and correlation analysis between DEGs and differentially accumulated flavonoids (DAFs). (a), genes related to flavonoid synthesis in 18 samples. LDOX, leucoanthocyanidin dioxygenase; F5H, cytochrome P450 84A1; 3'GT, anthocyanin 3'-O-beta-glucosyltransferase; AOMT, flavonoid 3',5'-methyltransferase; CYP75B1, flavonoid 3'-monooxygenase; FOMT, flavonoid 3'-O-methyltransferase; CHI, chalcone isomerase; CHS, chalcone synthase; COMT, caffeic acid 3-O-methyltransferase; FLS, flavonol synthase; ANR, anthocyanidin reductase. (b-d), correlation analysis between DEGs and DAFs of DK (b), DL (c), and AEX (d). ***, **, and * represent significant differences at P<0.001, P<0.010, and P<0.050 levels, respectively. The abbreviations are the same in the following figures. |
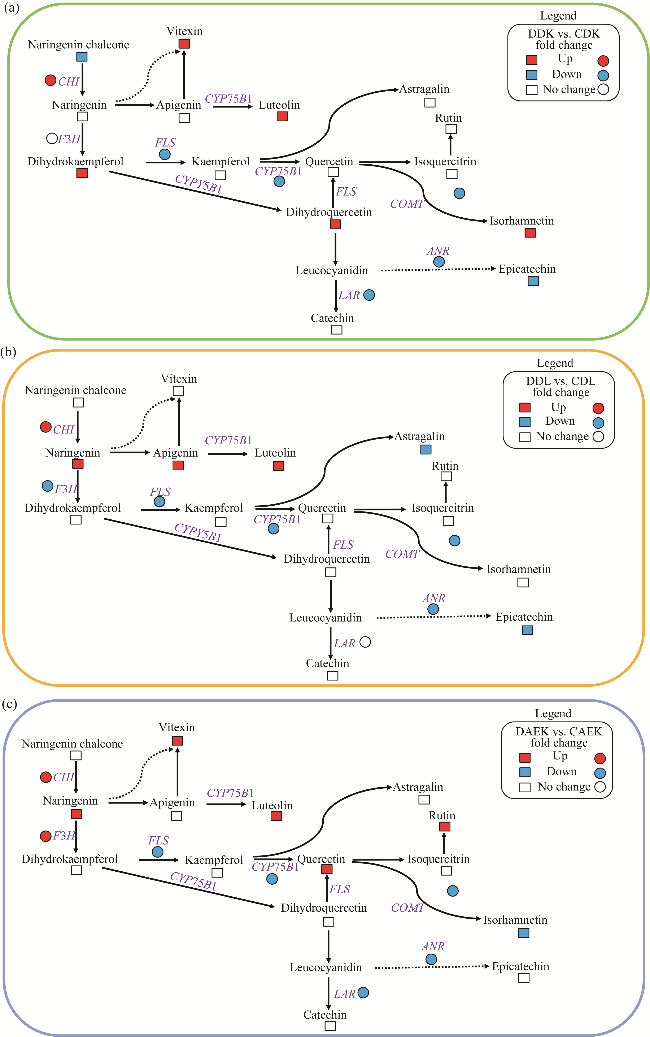
Fig. 5 Flavonoid biosynthesis pathway in response to drought stress in DK (a), DL (b), and AEX (c) ecotypes of sandrice. The heatmap colored red and blue indicates up-regulation and down-regulation of structural genes and flavonoid metabolites, respectively. The square and circle presented structural gene and flavonoid metabolites, respectively. |
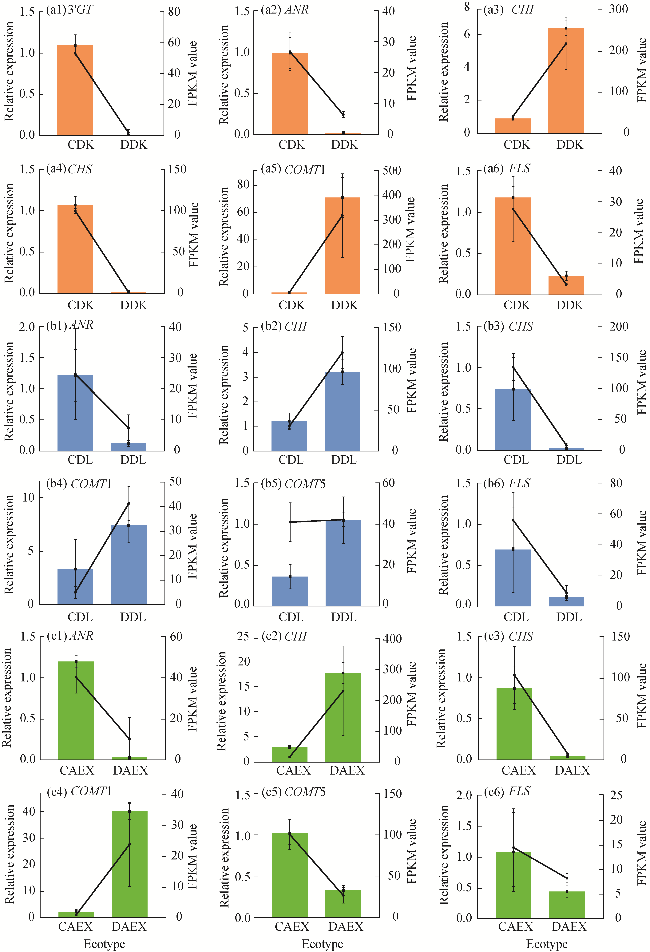
Fig. 6 Quantitative real-time polymerase chain reaction (qRT-PCR) validation of the transcriptome data in DK (a1-a6), DL (b1-b6) and AEX (c1-c6) ecotypes of sandrice. The line graph and the histogram with error bars present the relative expression calculated by the 2−ΔΔCt method and fragments per kilobase million (FPKM) value generated by ribonucleic acid sequence (RNA-Seq), respectively. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}