
Effect of long-term restoration on soil phosphorus transformation and desorption in the semi-arid degraded land, India
Received date: 2025-01-13
Revised date: 2025-04-22
Accepted date: 2025-05-09
Online published: 2025-08-13
Understanding how different vegetation-based restoration practices alter soil chemical and microbial characteristics is crucial, as restoration practices influence phosphorus (P) transformation and fractions and modify P adsorption behavior during the restoration process of degraded land. This study investigated the impacts of vegetation-based restoration practices on soil chemical and microbial parameters, P fractions, and patterns of P adsorption and desorption, and highlighted the combined influence on P availability. To evaluate the impact of vegetation-based restoration practices on P fractions and adsorption behavior in the semi-arid degraded land in India, this study compared three distinct tree-based restoration systems, including Leucaena leucocephala (Lam.) de Wit-based silviculture system (SCS), Acacia nilotica (L.) Willd. ex Delile-based silvopasture system (SPS), and Emblica officinalis Gaertn-based hortipasture system (HPS), with a natural grassland system (NGS) and a degraded fallow system (FS) as control. The soil samples across various soil depths (0-15, 15-30, and 30-45 cm) were collected. The findings demonstrated that SCS, SPS, and HPS significantly improved soil organic carbon (SOC) and nutrient availability. Moreover, SCS and SPS resulted in increased microbial biomass phosphorus (MBP) content and phosphatase enzyme activity. The P fractionation analysis revealed that ferrum-associated phosphorus (Fe-P) was the major P fraction, followed by aluminum-associated phosphorus (Al-P), reflecting the dominance of ferrum (Fe) and aluminum (Al) oxides in the semi-arid degraded land. Compared with FS, vegetation-based restoration practices significantly increased various P fractions across soil depths. Additionally, P adsorption and desorption analysis indicated a lower adsorption capacity in tree-based restoration systems than in FS, with FS soils adsorbing higher P quantities in the adsorption phase but releasing less P during the desorption phase. This study revealed that degraded soils responded positively to ecological restoration in terms of P fraction and desorption behavior, influencing the resupply of P in restoration systems. Consequently, litter rich N-fixing tree-based restoration systems (i.e., SCS and SPS) increased total phosphorus (TP) stock for plants and sustained the potential for long-term P supply in semi-arid ecosystems. With the widespread adoption of restoration practices across degraded landscapes, SCS and SPS would significantly contribute to soil restoration and improve productivity by maintaining the soil P supply in semi-arid ecosystems in India.
Jyotirmay ROY , Dipak Ranjan BISWAS , Biraj Bandhu BASAK , Ranjan BHATTACHARYYA , Shrila DAS , Sunanda BISWAS , Renu SINGH , Avijit GHOSH . Effect of long-term restoration on soil phosphorus transformation and desorption in the semi-arid degraded land, India[J]. Journal of Arid Land, 2025 , 17(6) : 846 -864 . DOI: 10.1007/s40333-025-0101-z
Table 1 Land system in the study area |
| Land system | Coverage area (hm2) | Sand (%) | Silt (%) | Clay (%) | Coordinate | Year of establishment | Soil erosion rate (Mg/(hm2•a)) |
|---|---|---|---|---|---|---|---|
| Silviculture system (SCS) | 3.0 | 53.80 | 30.70 | 15.50 | 25°31ʹ12ʺN, 78°32ʹ35ʺE | 2000 | 23.50 |
| Silvopasture system (SPS) | 1.1 | 59.80 | 24.70 | 15.50 | 25°31ʹ11ʺN, 78°32ʹ53ʺE | 2010 | 25.27 |
| Hortipasture system (HPS) | 1.2 | 51.80 | 34.70 | 13.50 | 25°31ʹ24ʺN, 78°32ʹ32ʺE | 1996 | 25.46 |
| Natural grassland system (NGS) | 2.0 | 59.80 | 24.70 | 15.50 | 25°31ʹ28ʺN, 78°32ʹ51ʺE | 2000 | 26.59 |
| Fallow system (FS) | 2.0 | 59.80 | 26.70 | 13.50 | 25°31ʹ14ʺN, 78°32ʹ52ʺE | 1980 | 53.29 |
Note: The soil erosion rate data were referred to Baradwal et al. (2023). |
Fig. 1 Landscape of land system in the semi-arid area in India. (a), Leucaena leucocephala (Lam.) de Wit based silviculture system (SCS); (b), Acacia nilotica (L.) Willd. ex Delile based silvopasture system (SPS); (c), Emblica officinalis Gaertn based hortipasture sytem (HPS); (d), natural grassland system (NGS); (e), fallow system (FS). |
Table 2 Phosphorus (P) fractionation scheme used by this study |
| P fraction | Extractant | Condition |
|---|---|---|
| Soluble and loosely bound phosphorus (Sal-P) | 1.000 M NH4Cl | Shake for 30 min and centrifuge at 10,000 r/min |
| Aluminum-associated phosphorus (Al-P) | 0.500 M NH4F | Shake for 17 h, centrifuge at 10,000 r/min, and wash with saturated NaCl |
| Ferrum-associated phosphorus (Fe-P) | 0.100 M NaOH | Shake for 1 h, centrifuge at 10,000 r/min, and wash with saturated NaCl |
| Calcium-associated phosphorus (Ca-P) | 0.250 M H2SO4 | Shake for 1 h, centrifuge at 10,000 r/min, and wash with saturated NaCl |
| Reductant soluble phosphorus (Res-P) | 0.300 M trisodium citrate dihydrate, 1.000 M NaHCO3, and 0.5 g Na2S2O4 | Water bath, stir, heat at 80°C, centrifuge at 10,000 r/min, and wash with saturated NaCl |
Table 3 Impact of land restoration on soil chemical property |
| Soil depth (cm) | Land system | pH | EC (dS/m) | SOC (g/kg) | AN (mg/kg) | AK (mg/kg) |
|---|---|---|---|---|---|---|
| 0-15 | SCS | 5.74±0.15a | 0.049±0.004a | 9.05±0.08a | 104.02±10.04a | 121.44±15.73a |
| SPS | 5.57±0.24a | 0.021±0.003b | 7.34±0.12ab | 96.82±6.23a | 35.26±8.70d | |
| HPS | 5.47±0.10a | 0.033±0.005bc | 5.93±0.06b | 83.86±4.19b | 60.82±5.55b | |
| NGS | 5.02±0.03b | 0.022±0.003bc | 5.33±0.02b | 83.85±4.42b | 54.91±4.35bc | |
| FS | 4.60±0.01b | 0.015±0.004c | 2.01±0.04c | 52.16±4.91c | 40.22±3.86c | |
| 15-30 | SCS | 5.90±0.00a | 0.044±0.004a | 5.13±0.05a | 82.19±8.15a | 56.32±8.72a |
| SPS | 5.57±0.16ab | 0.021±0.003a | 4.02±0.06b | 82.19±8.27a | 25.26±3.55c | |
| HPS | 5.54±0.19ab | 0.036±0.006b | 3.67±0.04b | 63.42±6.06b | 32.16±6.94bc | |
| NGS | 5.36±0.13bc | 0.020±0.003b | 2.66±0.01c | 69.29±4.28b | 41.04±8.11b | |
| FS | 4.99±0.15c | 0.019±0.004b | 1.56±0.02c | 42.95±2.83c | 33.92±4.03bc | |
| 30-45 | SCS | 5.82±0.05a | 0.042±0.005a | 4.47±0.08a | 74.48±4.91a | 51.83±6.86a |
| SPS | 5.58±0.15a | 0.020±0.001b | 4.52±0.06ab | 74.50±4.22a | 22.33±4.89c | |
| HPS | 5.58±0.02a | 0.029±0.003b | 3.47±0.04b | 59.76±4.24b | 29.54±4.76bc | |
| NGS | 5.25±0.01b | 0.021±0.003b | 2.36±0.02c | 59.72±4.17b | 44.11±5.33b | |
| FS | 4.90±0.05c | 0.020±0.006b | 1.61±0.01c | 39.29±3.19c | 32.12±5.06bc |
Note: EC, electrical conductivity; SOC, soil organic carbon; AN, available nitrogen; AK, available potassium. Different lowercase letters within the same soil depth indicate statistical significant differences among different land systems at P<0.05 level. Mean±standard deviation (SD). |
Table 4 Impact of land restoration practice on soil microbial property |
| Microbial parameter | Soil depth (cm) | Land system | ||||
|---|---|---|---|---|---|---|
| SCS | SPS | HPS | NGS | FS | ||
| DHA activity (μg TPF/(g•24h)) | 0-15 | 78.42±3.33a | 46.33±4.15b | 45.91±5.66b | 50.52±1.64b | 31.62±3.12c |
| 15-30 | 50.82±3.06a | 32.79±2.49b | 26.86±2.63b | 30.85±0.62b | 25.54±0.97b | |
| 30-45 | 21.53±1.50a | 18.18±0.39b | 17.57±0.62b | 18.08±1.32b | 16.35±0.27b | |
| ACP activity (μg PNP/(g•h)) | 0-15 | 146.50±1.84a | 120.35±2.21b | 89.29±1.93c | 74.80±7.13d | 46.51±5.06e |
| 15-30 | 94.42±4.05a | 75.44±5.29ab | 85.62±2.60b | 39.08±2.14c | 22.71±0.43d | |
| 30-45 | 90.44±2.52a | 55.61±8.61b | 68.55±5.74b | 28.04±4.86c | 15.30±0.52c | |
| ALP activity (μg PNP/(g•h)) | 0-15 | 75.31±5.34a | 42.22±1.41b | 43.11±4.71b | 21.38±1.03c | 15.60±2.17c |
| 15-30 | 67.12±2.46a | 37.46±1.45b | 35.72±1.54b | 10.85±1.44c | 10.4±2.17c | |
| 30-45 | 44.06±7.41a | 33.53±6.51a | 29.37±2.35a | 10.60±0.61b | 9.6±1.44b | |
| MBC (mg/kg) | 0-15 | 928.48±24.21a | 805.29±40.50ab | 831.83±16.15ab | 683.46±101.25b | 428.49±68.33c |
| 15-30 | 849.17±36.79a | 743.62±26.48a | 782.10±35.39a | 602.84±37.40b | 378.45±66.48c | |
| 30-45 | 637.33±22.10a | 512.32±36.22b | 540.46±18.74b | 378.92±38.35c | 348.35±43.36c | |
| MBP (mg/kg) | 0-15 | 8.80±0.67a | 8.16±0.61a | 5.59±0.46b | 4.93±0.21b | 4.43±0.20b |
| 15-30 | 6.26±0.47a | 4.15±0.53b | 4.26±0.23b | 4.16±0.60b | 3.18±0.41b | |
| 30-45 | 4.41±0.27a | 3.83±0.08ab | 3.53±0.44ab | 3.47±0.80ab | 2.88±0.20b | |
Note: DHA, dehydrogenase; ACP, acid phosphatase; ALP, alkaline phosphatase; MBC, microbial biomass carbon; MBP, microbial biomass phosphorus. Mean±SD. Different lowercase letters within the same soil depth indicate statistical significant differences among different land systems at P<0.05 level. |
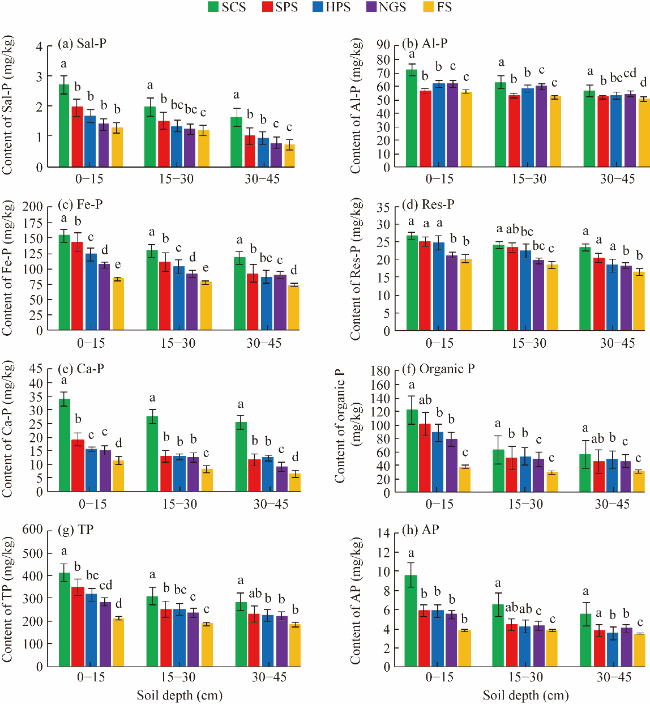
Fig. 2 Impact of different land restoration practices on soil phosphorus (P) fraction at different soil depths. (a), soluble and loosely bound-phosphorus (Sal-P); (b), aluminium-associated phosphorus (Al-P); (c), ferrum-associated phosphorus (Fe-P); (d), calcium-associated phosphorus (Ca-P); (e), reductant soluble phosphorus (Res-P); (f), organic P; (g), total phosphorus (TP); (h), available phosphorus (AP). Different lowercase letters within the same soil depth indicate statistical significant differences among different land systems at P<0.05 level. Bars represent standard errors. |
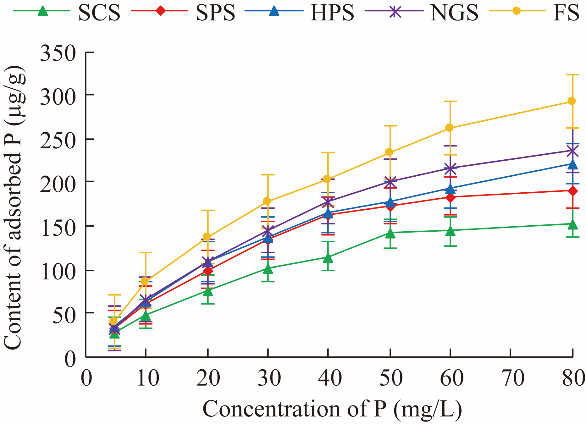
Fig. 3 P adsorption curve under different restoration practices. Bars represent standard errors. |
Table 5 Langmuir and Freundlich adsorption parameter for different land systems |
| Land system | Langmuir adsorption parameter | Freundlich adsorption parameter | |||
|---|---|---|---|---|---|
| b (µg/g) | k (mL/µg) | y (mL/g) | a | n | |
| SCS | 190±10b | 0.069±0.013b | 12.9±1.6b | 21±1c | 1.95±0.05b |
| SPS | 231±21ab | 0.093±0.013b | 21.0±1.2b | 29±3bc | 1.98±0.10b |
| HPS | 271±44ab | 0.085±0.003b | 22.8±2.8b | 34±3b | 2.01±0.07b |
| NGS | 294±55ab | 0.081±0.015b | 22.6±2.6b | 36±5b | 2.04±0.02b |
| FS | 315±30a | 0.144±0.013a | 45.3±5.4a | 62±8a | 2.43±0.04a |
Note: b, maximum phosphorus adsorption capacity; k, phosphorus binding affinity; y, maximum phosphorus buffering capacity; a, number of phosphorus adsorption sites; n, phosphorus bonding energy. Mean±SD. Different lowercase letters within the same parameter indicate statistical significant differences among different land systems at P<0.05 level. |
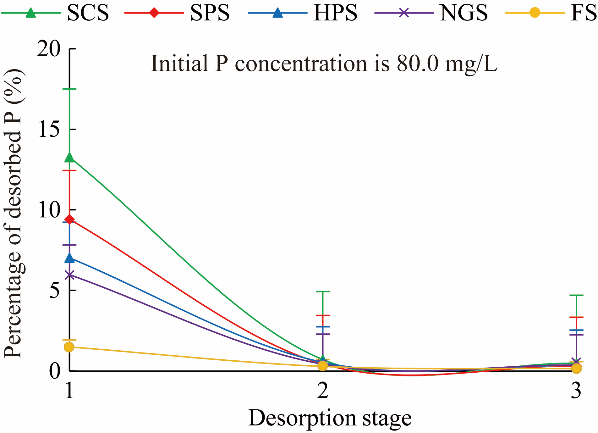
Fig. 4 P desorption curve under different restoration practices. Bars represent standard errors. |
Table 6 Aboveground biomass, litterfall, and root biomass in different land systems |
| Land system | Aboveground biomass (Mg/hm2) | Litterfall (g/m2) | Root biomass (g/m2) | ||
|---|---|---|---|---|---|
| 0-15 cm | 15-30 cm | 30-45 cm | |||
| SCS | 12.53±1.24a | 147.44±11.02b | 84.67±7.22a | 71.37±7.12a | 34.69±3.11b |
| SPS | 9.89±0.97b | 126.25±12.64c | 58.64±5.42b | 75.67±7.31a | 54.27±5.22a |
| HPS | 10.24±0.87b | 198.14±8.47a | 40.33±4.02c | 62.34±6.47b | 52.21±5.17a |
| NGS | 3.25±0.35c | 57.14±3.29d | 38.33±3.02c | 32.34±3.17c | 25.21±2.17c |
| FS | 0.03±0.00d | 15.2.14±3.28e | 5.28±0.52d | 4.67±0.45d | 1.27±0.22d |
Note: Mean±SD. Different lowercase letters within the same variable indicate statistical significant differences among different land systems at P<0.05 level. |
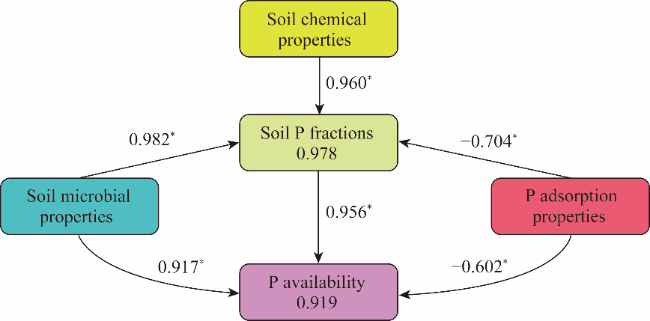
Fig. 5 Structural equation model illustrating the influence of microbial, soil chemical, and P adsorption properties on the distribution of soil P fractions and P availability. The numbers in frames represent the proportion of variance explained for the respective latent variable; while the numbers above arrows represent z-standardized path coefficients, of which, positive numbers represent positive relationship between two variables and negative numbers represent negative relationship between two variables. *, P<0.05 level. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}