
Seed germination response of the invasive Haloxylon persicum in Tunisia
Received date: 2024-07-04
Revised date: 2024-11-06
Accepted date: 2024-11-14
Online published: 2025-08-13
Biological invasion represents a major worldwide threat to native biodiversity and environmental stability. Haloxylon persicum was introduced to Tunisia (North Africa) with Saharan bioclimate in 1969 to fix sandy dunes. Since then, it has gained significant interest for its potential to colonize, proliferate, and become naturalized in Tunisia. Hence, understanding the seed germination response of H. persicum to abiotic conditions, including temperature, water stress, and salt stress, is crucial for predicting its future spread and adopting effective control strategies. Our work investigated the germination behavior of this invasive plant species by incubation at temperatures from 10.0°C to 35.0°C and at various osmotic potentials (-2.00, -1.60, -1.00, -0.50, and 0.00 MPa) of polyethylene glycol-6000 (PEG6000, indicating water stress) and sodium chloride (NaCl, indicating salt stress) solutions. Results showed remarkable correlations among the seed functional traits of H. persicum, indicating adaptive responses to local environmental constraints. The maximum germination rate was recorded at 25.0°C with a rate of 0.39/d. Using the thermal time model, the base temperature was recorded at 8.4°C, the optimal temperature was 25.5°C, and the ceiling temperature was found at 58.3°C. Besides, based on the hydrotime model, the base water potential showed lower values of -7.74 and -10.90 MPa at the optimal temperatures of 25.0°C and 30.0°C, respectively. Also, the species was found to have excellent tolerance to drought (water stress) compared to salt stress, which has implications for its potential growth into new habitats under climate change. Combining ecological and physiological approaches, this work elucidates the invasive potential of H. persicum and contributes to the protection of species distribution in Tunisian ecosystems.
Hanen FARHAT , Khouloud KRICHEN , Khouloud ZAGOUB , Mohamed CHAIEB . Seed germination response of the invasive Haloxylon persicum in Tunisia[J]. Journal of Arid Land, 2024 , 16(12) : 1744 -1759 . DOI: 10.1007/s40333-024-0065-4
Table 1 Seed functional traits of Haloxylon persicum |
| Seed functional trait | Mean±SE | Coefficient of variation (%) |
|---|---|---|
| Thousand seed weight (g) | 3.15±0.00 | 0.95 |
| Percentage of embryoless seeds (%) | 22.51±1.68 | 45.50 |
| Embryo viability (%) | 74.33±0.00 | 15.71 |
| Seed diameter (mm) | 2.00±0.00 | 0.20 |
| Seed area (mm2) | 3.14±0.07 | 0.02 |
| Seed water content (%) | 14.76±0.06 | 0.03 |
| Seed shape | Round | |
| Seed color | Brownish grey | |
| Embryo form | Spirally twisted |
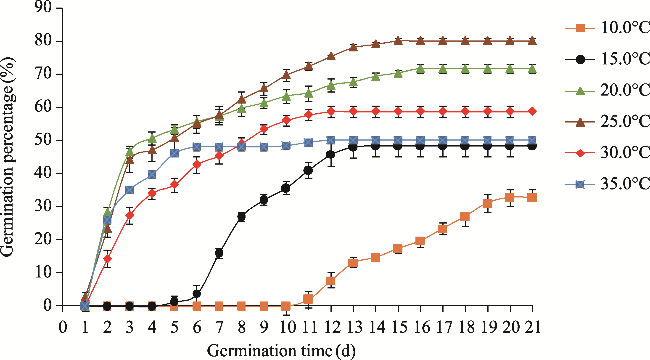
Fig. 1 Germination percentage of Haloxylon persicum at different temperatures. Each point represents the mean of replicates of 50 seeds. Bars mean standard deviations. |
Table 2 Seed germination parameters of H. persicum at different temperatures |
| Temperature (°C) | Final germination percentage (%) | Initial germination time (d) | Final germination time (d) | Mean germination time (d) |
|---|---|---|---|---|
| 10.0 | 33.00±2.42b | 12.00±0.41a | 20.00±0.41c | 9.36±0.85a |
| 15.0 | 48.00±3.04c | 6.00±1.26c | 13.00±0.41b | 13.18±0.56b |
| 20.0 | 72.00±1.17a | 2.00±0.00a | 15.00±1.51b | 10.25±0.24a |
| 25.0 | 80.00±0.09a | 1.50±0.54a | 14.00±0.99a | 10.14±0.55a |
| 30.0 | 51.00±1.63b | 2.00±0.00b | 10.00±1.22b | 10.35±0.56a |
| 35.0 | 50.00±1.26b | 2.00 ±0.00b | 11.00±0.75c | 10.73±0.22b |
Note: Mean±SD. Different lowercase letters in the same column indicate significant differences among different temperatures at P<0.05 level. |
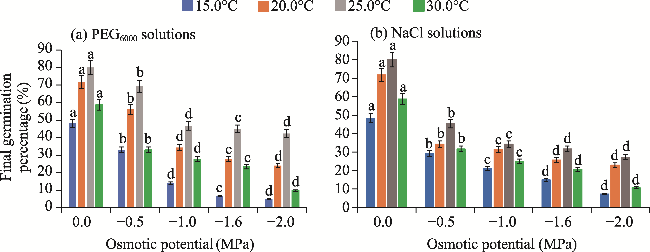
Fig. 2 Effect of osmotic potentials of PEG6000 (polyethylene glycol-6000; a) and NaCl (sodium chloride; b) on the seed germination of H. persicum. Different lowercase letters indicate significant differences of final germination percentage at P<0.05 level. Bars mean standard deviations. |
Table 3 Effects of PEG6000 (polyethylene glycol-6000) and NaCl (sodium chloride) and their interactions with temperature on the final germination percentage of H. persicum based on the two-way analysis of variance (ANOVA) |
| Salt stress | Water stress | |||||
|---|---|---|---|---|---|---|
| Temperature | NaCl | Temperature×NaCl | Temperature | PEG6000 | Temperature×PEG6000 | |
| df | 1.39 | 2.24 | 2.68 | 2.15 | 1.16 | 3.96 |
| F statistic | 1571.77 | 153.93 | 44.62 | 909.27 | 255.09 | 26.58 |
| P-value | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
Note: df, degrees of freedom. |
Table 4 Estimated hydrotime model parameters describing the seed germination of H. persicum at different temperatures under the water stress (PEG6000 solutions) and salt stress (NaCl solutions) treatments |
| Temperature (°C) | Water stress | Salt stress | ||||||
|---|---|---|---|---|---|---|---|---|
| Ψb(50) (MPa) | θH (MPa•d) | σΨb(g) (MPa) | R2 | Ψb(50) (MPa) | θH (MPa•d) | σΨb(g) (MPa) | R2 | |
| 15.0 | -4.14 | 37.71 | 1.26 | 0.76 | -3.61 | 14.50 | 2.07 | 0.71 |
| 20.0 | -7.08 | 29.60 | 3.97 | 0.61 | -7.03 | 25.84 | 4.02 | 0.61 |
| 25.0 | -7.74 | 22.86 | 4.75 | 0.57 | -4.66 | 21.09 | 2.48 | 0.48 |
| 30.0 | -10.90 | 47.06 | 5.63 | 0.63 | -6.66 | 20.30 | 3.47 | 0.60 |
Note: Ψb(50), base water potential when the germination percentage is 50.00%; θH, hydrotime constant; σΨb(g), standard deviation of the base water potential at a specific germination percentage; R², coefficient of determination. |
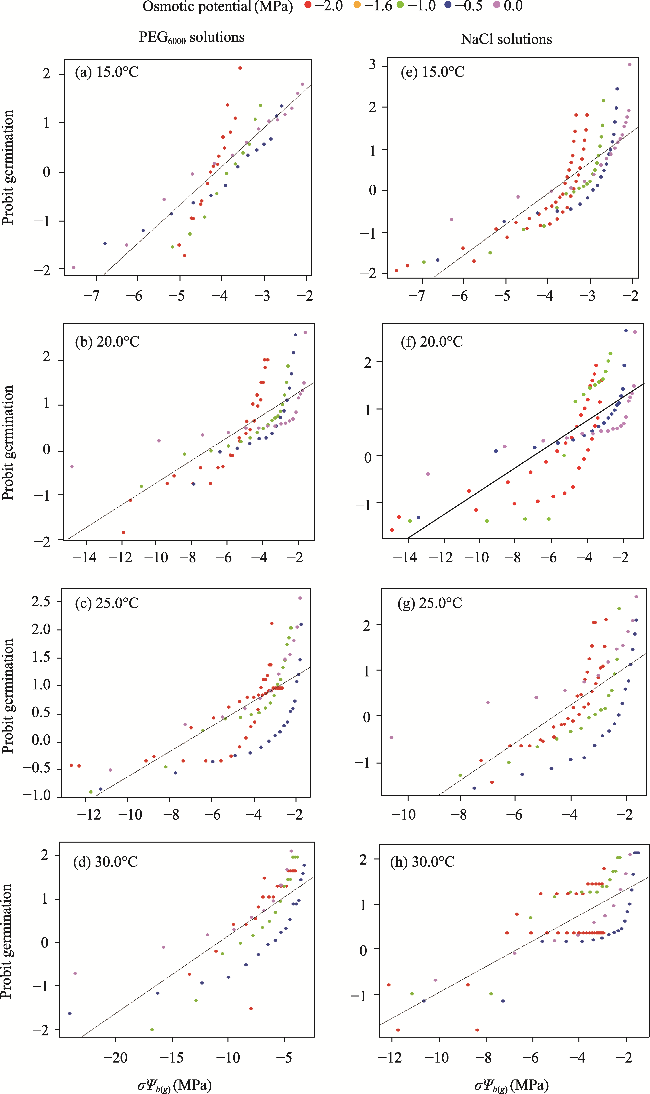
Fig. 3 Relationship between the probit germination model of H. persicum and σΨb(g) at different osmotic potentials of PEG6000 (a-d) and NaCl (e-h) solutions at different temperatures (15.0°C, 20.0°C, 25.0°C and 30.0°C). σΨb(g), standard deviation of the base water potential defined at a specific germination percentage. |
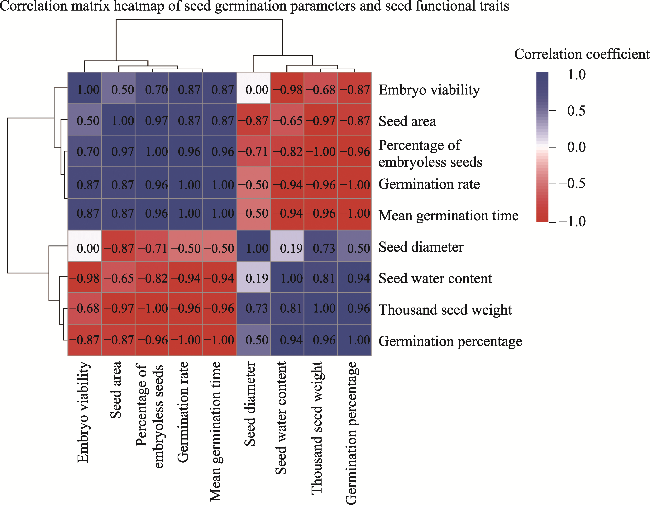
Fig. 4 Heat map correlation between seed germination parameters and seed functional traits of H. persicum at 25.0°C |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
ISTA (International Seed Testing Association). 2013. International Rules for Seed Testing. Wallisellen: ISTA.
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}