
Impact of trimming intensity on the growth of mangrove in Iran
Received date: 2024-06-10
Revised date: 2024-11-06
Accepted date: 2024-11-30
Online published: 2025-08-13
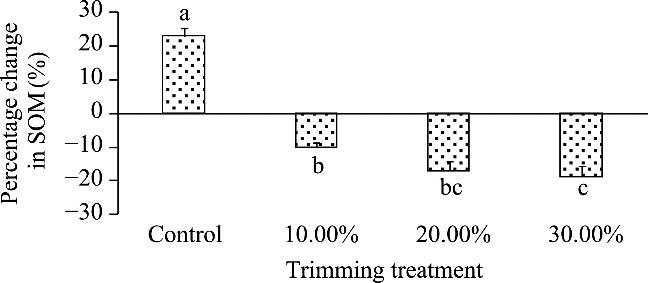
Mangrove forests in southern Iran are of high ecological and economic importance. These forests are being threatened because of uncontrolled harvesting to provide fodder for livestock. The objective of this study is to provide recommendations for appropriate harvesting intensities by quantifying the effect of different harvesting intensities on vegetative and vigor characteristics of mangrove trees. This study was conducted using a randomized complete block design comprising four treatments (10.00%, 20.00%, and 30.00% trimming, along with a control) replicated three times. Vegetative characteristics were measured before and after trimming (five-year period) and analyzed using generalized linear model statistical analysis. The growths of the average diameter of canopy, canopy area, canopy volume, canopy height, tree height, and collar diameter in the control treatment were all significantly higher than those in the trimming treatments. In addition, there was a decreasing trend in leaf fresh and dry mass, leaf area index, total area of canopy leaves, and health status of tree in the trimming treatments. For example, the percentage change in fresh and dry leaf mass in the control treatment was positive (29.87% and 38.31%, respectively), whereas the trimming treatments of 10.00%, 20.00% and 30.00% had negative effects (-7.01% and -4.79%, -11.32% and -14.30%, and -15.84% and -17.29%, respectively). In addition, the changes in leaf area index in the control (4.95%) and 30.00% trimming (-24.57%) treatments were the highest and lowest, respectively. The percentage change in soil organic matter in the control, 10.00%, 20.00%, and 30.00% treatments were 22.94%, -9.90%, -16.91%, and -18.68%, respectively. The study demonstrated that gray mangrove trees were highly sensitive to canopy trimming, with even minimal trimming intensities negatively affecting vegetative growth and soil organic matter. Therefore, it is recommended that cutting and trimming of mangrove trees should be prevented even at low intensity to preserve mangrove ecosystem health and resilience against environmental stressors.
Maryam MOSLEHI , Akram AHMADI , Tom PYPKER . Impact of trimming intensity on the growth of mangrove in Iran[J]. Journal of Arid Land, 2024 , 16(12) : 1701 -1713 . DOI: 10.1007/s40333-024-0111-2
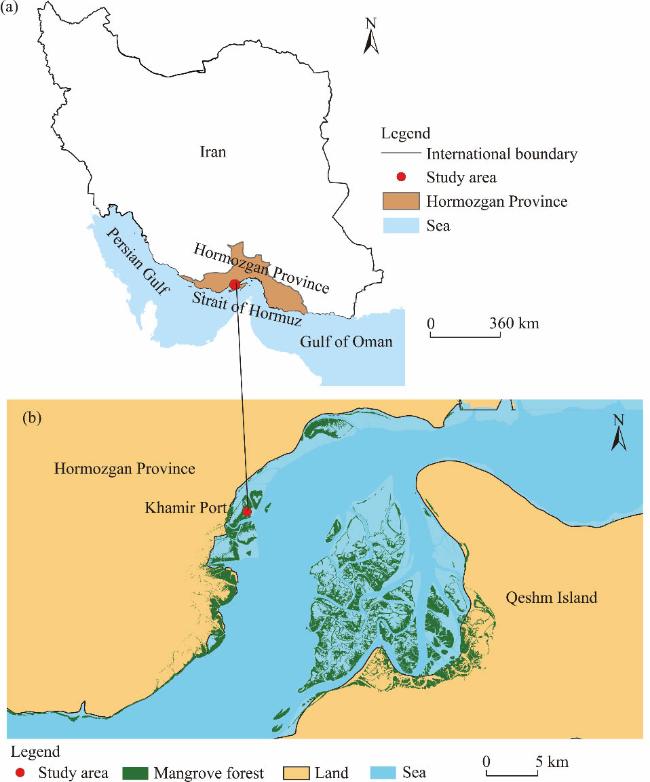
Fig. 1 Location of the study area (a) and the distribution range of mangrove forests (b) in Khamir Port, Iran |
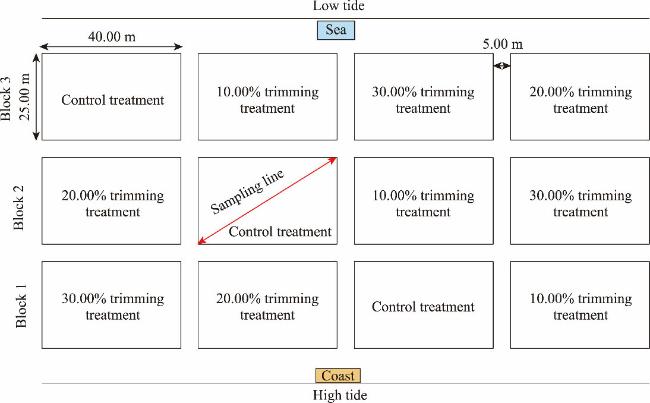
Fig. 2 Schematic layout of sampling |
Table 1 Rubric for assessing the grade of tree health |
| Grade | Score | Health indicator | |||
|---|---|---|---|---|---|
| Color of leaf | Shape of canopy | Density of canopy | Number of dry microbranches | ||
| Healthy | 5 | Bright green | Symmetrical and healthy | Dense | <10 |
| Moderate | 3 | Pale green | Relatively symmetrical and average size | Semi-dense | 10-30 |
| Weak | 1 | Yellow or brown | Asymmetrical, small, or damaged | Very sparse | >30 |
Table 2 Analysis of variance for the growth of mangroves across various treatments |
| Variable | Source of variable | df | Mean square | Fisher coefficient |
|---|---|---|---|---|
| Small diameter of canopy | Block | 2 | 0.085 | 6.23* |
| Treatment | 3 | 0.363 | 26.70** | |
| Error | 6 | 0.014 | ||
| Large diameter of canopy | Block | 2 | 0.267 | 12.40** |
| Treatment | 3 | 0.293 | 13.63** | |
| Error | 6 | 0.022 | ||
| Average diameter of canopy | Block | 2 | 0.160 | 10.57* |
| Treatment | 3 | 0.320 | 21.16** | |
| Error | 6 | 0.010 | ||
| Canopy area | Block | 2 | 25.670 | 28.71** |
| Treatment | 3 | 42.230 | 47.24** | |
| Error | 6 | 0.890 | ||
| Canopy volume | Block | 2 | 4415.840 | 12.73** |
| Treatment | 3 | 7207.430 | 20.77** | |
| Error | 6 | 346.880 | ||
| Canopy height | Block | 2 | 0.003 | 7.06* |
| Treatment | 3 | 0.017 | 38.37** | |
| Error | 6 | 0.000 | ||
| Tree height | Block | 2 | 0.004 | 7.21* |
| Treatment | 3 | 0.028 | 57.70** | |
| Error | 6 | 0.000 | ||
| Collar diameter | Block | 2 | 0.020 | 0.61ns |
| Treatment | 3 | 2.620 | 78.86** | |
| Error | 6 | 0.033 |
Note: df, degree of freedom; *, significant difference at P<0.05 level; **, significant difference at P<0.01 level; ns, no significant difference. |
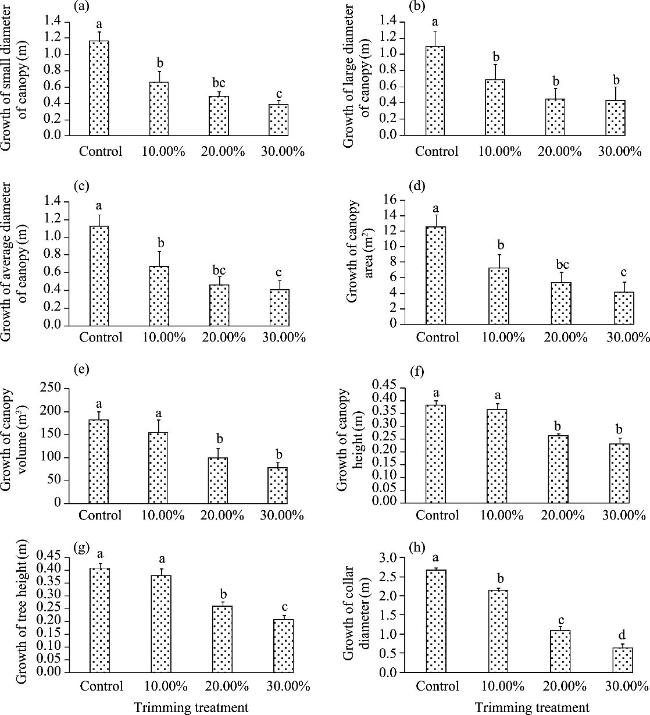
Fig. 3 Growth of mangroves across various treatments over a five-year period. (a), growth of small diameter of canopy; (b), growth of large diameter of canopy; (c), growth of average diameter of canopy; (d), growth of canopy area; (e), growth of canopy volume; (f), growth of canopy height; (g), growth of tree height; (i), growth of collar diameter. Different lowercase letters indicate significant differences among different treatments at P<0.05 level; and bar represents standard deviation. |
Table 3 Analysis of variance for percentage changes in leaf characteristics and health status of tree across various treatments |
| Variable | Source of variable | df | Mean square | Fisher coefficient |
|---|---|---|---|---|
| Leaf fresh mass (LFM) | Block | 2 | 22.190 | 0.64ns |
| Treatment | 3 | 1316.560 | 38.27** | |
| Error | 6 | 34.390 | ||
| Leaf dry mass (LDM) | Block | 2 | 20.450 | 0.58ns |
| Treatment | 3 | 1993.890 | 57.02** | |
| Error | 6 | 34.960 | ||
| Leaf area index (LAI) | Block | 2 | 40.250 | 2.56ns |
| Treatment | 3 | 570.640 | 69.70** | |
| Error | 6 | 27.920 | ||
| Total area of canopy leaves (TAL) | Block | 2 | 82.370 | 3.14ns |
| Treatment | 3 | 2251.250 | 64.56** | |
| Error | 6 | 32.290 | ||
| Health status of tree | Block | 2 | 167.990 | 1.44ns |
| Treatment | 3 | 530.640 | 20.43** | |
| Error | 6 | 36.960 |
Note: **, significant difference at P<0.01 level; ns, no significant difference. |
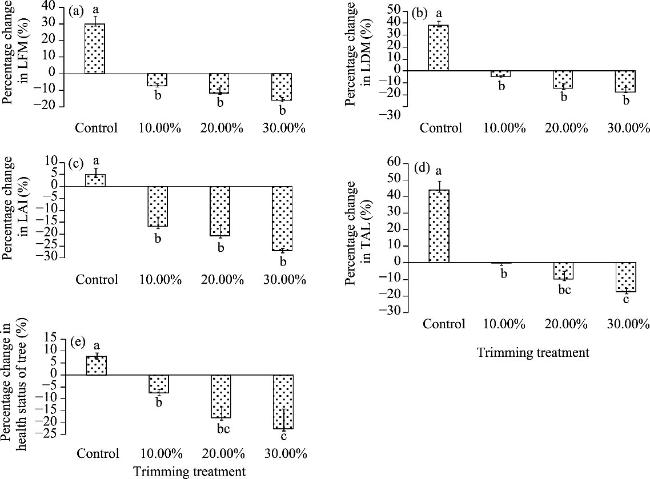
Fig. 4 Comparison of percentage change in leaf characteristics and health status of tree across various treatments over a five-year period using Duncan's multiple range test. (a), leaf fresh mass (LFM); (b), leaf dry mass (LDM); (c), leaf area index (LAI); (d), total area of canopy leaves (TAL); (e), health status of tree. Different lowercase letters indicate significant differences among different treatments at P<0.05 level; and bar represents standard deviation. |
Table 4 Analysis of variance of the percentage change in soil organic matter (SOM) across various treatments |
| Variable | Source of variable | df | Mean square | Fisher coefficient |
|---|---|---|---|---|
| SOM | Block | 2 | 22.960 | 1.32ns |
| Treatment | 3 | 11.320 | 64.72** | |
| Error | 6 | 17.490 |
Note: **, significant difference at P<0.01 level; ns, no significant difference. |
Fig. 5 Comparison of percentage change in soil organic matter (SOM) across various treatments over a five-year period using Duncan's multiple range test. Different lowercase letters indicate significant differences among different treatments at P<0.05 level; and bar represents standard deviation. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
FAO (Food and Agricultural Organization). 2003. Status and Trends in Mangrove Area Extent World-wide. [2024-05-18]. http://www.fao.org/4/j1533e/J1533E01.htm.
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}