
Effects of temperature and salinity on the seed germination of Limonium strictissimum (Salzm.) Arrigoni in the Mediterranean Basin
Received date: 2024-08-09
Revised date: 2024-10-30
Accepted date: 2024-11-26
Online published: 2025-08-12
Salt stress might be an important factor that decreases the emergence of seedlings and reduces plants' growth, causing their endangered status. However, the effects of salt stress on the germination of the Mediterranean species are less concern. Limonium strictissimum (Salzm.) Arrigoni, part of the group related to Limonium articulatum (Loisel.) Kuntze, is a Cyrno-Sardinian endemism, which is included in the International Union for Conservation of Nature (IUCN) Red Lists as Endangered (EN). Fresh seeds of L. strictissimum harvested from the only known population in Sardinia and a population in southeastern Corsica were used with the aims to study the effects of temperature and salinity on the seed germination and to evaluate the ability to recover their germination after exposure to salt stress. In both populations, empty fruits with a lower percentage were observed in Sardinia than in Corsica. The seeds showed a high germination capacity, which was not influenced by temperature and did not even differ between the two populations. Compared with non-saline condition, the presence of salt in the substrate, independently of the concentration tested, inhibited seed germination in both populations, with germination percentages never exceeding 40%. The Sardinian population showed a recovery capacity after exposure to high salt stress, always above 50% regardless of the salt concentration and incubation temperature considered. The seeds previously exposed to 125 mM NaCl at 30°C had the lowest recovery percentage (56%), while the highest recovery percentage (84%) was recorded at 25°C with a concentration of 250 mM NaCl. Differently, the Corsican population recorded a higher recovery percentage (54%) in seeds previously exposed to 500 mM NaCl at 20°C, while for the concentration of 125 mM NaCl, the best recovery percentage (11%) was recorded at 30°C. The ability of L. strictissimum to recover germination after salt exposure can be interpreted as an adaptation to the coastal habitats in which it grows. This study provides new insights into the ecophysiology of L. strictissimum seed germination, which could help preserve and implement effective conservation measures for this endangered species with restricted populations.
Ludovica DESSÌ , Alba CUENA-LOMBRAÑA , Lina PODDA , Marco PORCEDDU , Mauro FOIS , Lillia FAUSTI , Carole PIAZZA , Gianluigi BACCHETTA . Effects of temperature and salinity on the seed germination of Limonium strictissimum (Salzm.) Arrigoni in the Mediterranean Basin[J]. Journal of Arid Land, 2025 , 17(2) : 259 -270 . DOI: 10.1007/s40333-025-0004-z
Table 1 Information of Limonium strictissimum (Salzm.) Arrigoni investigated in this study |
| Population | Locality | Island (Country) | Coordinates | Substrate | Date of collecting (dd/mm/yyyy) | Number of collected seeds |
|---|---|---|---|---|---|---|
| Sardinian population | Punta Rossa | Sardinia (Italy) | 41°10′11″N 09°28′13″E | Granites | 14/10/2022 | 10,795 |
| Corsican population | Plage di Maora | Corsica (France) | 41°24′28″N 09°12′59″E | Gabbros | 14/09/2022 | 990 |
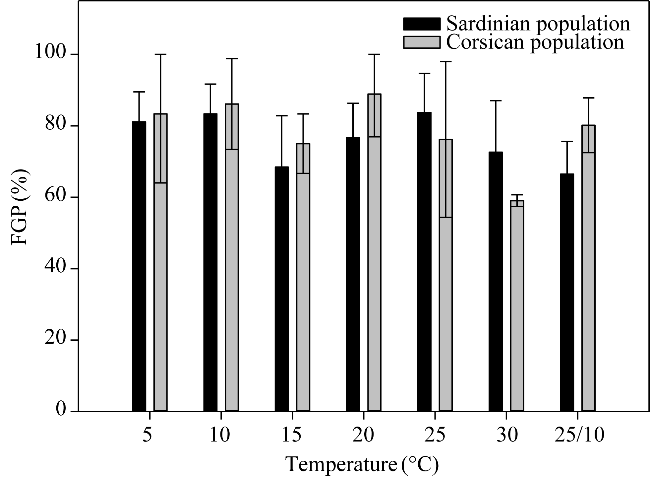
Fig. 1 Final germination percentage (FGP) of Limonium strictissimum (Salzm.) Arrigoni seeds from different populations incubated at constant (5°C, 10°C, 15°C, 20°C, 25°C, and 30°C) and alternating (25°C/10°C) temperatures. Bars are standard errors. |
Table 2 Generalised linear model (GLM) result of final germination percentage (FGP) in L. strictissimum seeds under different temperatures and populations |
| Factor | df | Sum square | Mean square | F | P value |
|---|---|---|---|---|---|
| Population (P) | 1 | 0.005 | 0.005 | 0.281 | 0.600 |
| Temperature (T) | 6 | 0.177 | 0.029 | 1.464 | 0.226 |
| P×T | 6 | 0.089 | 0.014 | 0.737 | 0.624 |
Table 3 GLM result of FGP in L. strictissimum seeds under different salt stresses, temperatures, and populations |
| Factor | df | Sum square | Mean square | F | P value |
|---|---|---|---|---|---|
| Salt stress | 3 | 6.917 | 2.305 | 318.856 | <0.001*** |
| Temperature | 2 | 0.073 | 0.036 | 5.031 | <0.050* |
| Population | 1 | 0.007 | 0.007 | 0.992 | 0.324 |
Note: *, P<0.050 level; ***, P<0.001 level. |
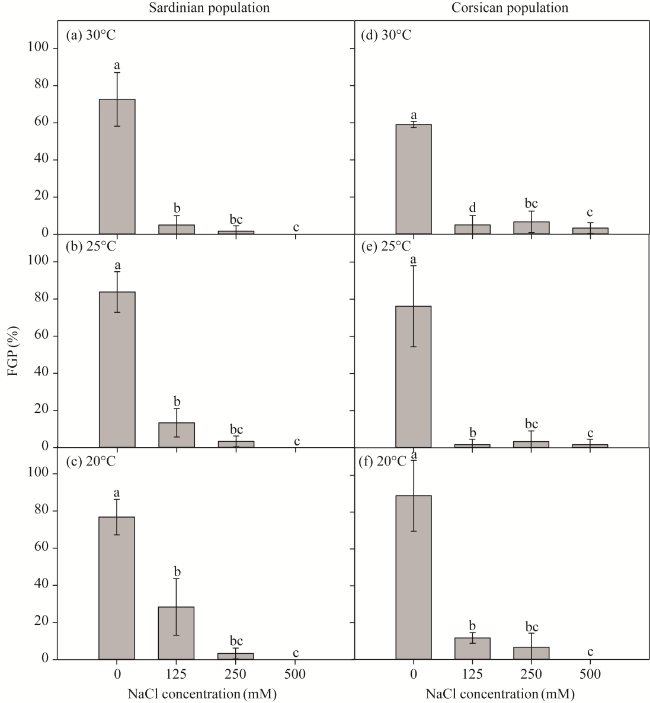
Fig. 2 Effect of different concentrations of NaCl (0, 125, 250, and 500 mM) on FGP of L. strictissimum incubated at constant temperatures (20°C, 25°C, and 30°C) for different populations. Different lowercase letters indicate significant differences among different NaCl concentrations at P<0.050 level. (a-c), 30°C, 25°C, and 20°C in the Sardinian population; (d-f), 30°C, 25°C, and 20°C in the Corsican population. Bars are standard errors. |
Table 4 GLM result of recovery percentage (RP) in L. strictissimum seeds under different salt stresses, temperatures, populations, and their interactions |
| Factor | df | Sum square | Mean square | F | P value |
|---|---|---|---|---|---|
| Salt stress (S) | 2 | 0.938 | 0.469 | 11.682 | <0.001*** |
| Temperature (T) | 2 | 0.034 | 0.017 | 0.426 | 0.656 |
| Population (P) | 1 | 1.780 | 1.780 | 44.314 | <0.001*** |
| S×T | 4 | 0.081 | 0.020 | 0.505 | 0.732 |
| S×P | 2 | 0.352 | 0.176 | 4.389 | <0.050* |
| T×P | 2 | 0.039 | 0.019 | 0.492 | 0.615 |
| S×T×P | 4 | 0.172 | 0.042 | 1.059 | 0.390 |
Note: *, P<0.050 level; ***, P<0.001 level. |
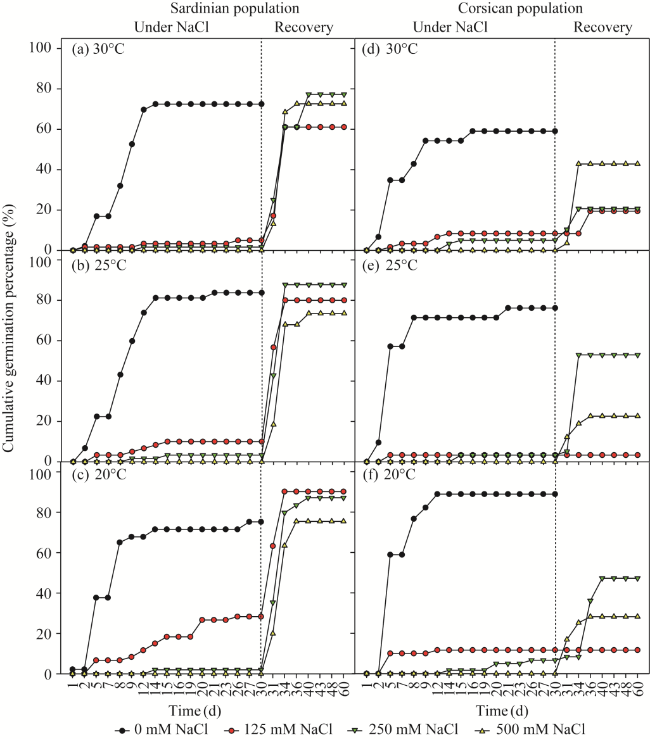
Fig. 3 Cumulative germination percentage of L. strictissimum seeds under different temperatures (20°C, 25°C, and 30°C) and salt stresses (0, 125, 250, and 500 mM NaCl) for different populations. (a-c), 30°C, 25°C, and 20°C in the Sardinian population; (d-f), 30°C, 25°C, and 20°C in the Corsican population. The vertical line indicates the beginning of the recovery phase. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
ISTA (International Seed Testing Association). 2006. International Rules for Seed Testing. [2024-04-17].
|
| [23] |
IUCN (International Union for Conservation of Nature). 2024. The National Red List Database. [2024-02-08].
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
R Development Core Team. 2023. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. [2024-02-04]. https://www.rproject.org/
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}