
Applying joint species distribution modelling to assess the relative influence of ecological filters on community assembly in the El Bayadh steppe, Algeria
Received date: 2025-01-02
Revised date: 2025-03-29
Accepted date: 2025-04-03
Online published: 2025-08-12
Understanding plant community assembly is crucial for effective ecosystem conservation and restoration. The ecological filter framework describes community assembly as a process shaped by dispersal, environmental, and biotic filters. Additionally, functional traits and phylogenetic relationships are increasingly recognized as important factors influencing species coexistence and community structure. However, both the ecological filter framework and the roles of functional traits and phylogeny in community assembly remain underexplored in the Algerian steppes—particularly in the El Bayadh region, where ongoing vegetation degradation threatens ecosystem stability. This study applied Hierarchical Modeling of Species Communities (HMSC) as an integrative approach to assess how ecological filters influence plant community assembly in the El Bayadh steppe and to evaluate the roles of functional traits and phylogenetic relationships in this process. Environmental data—including soil properties, topography, precipitation, and land use types (grazing and exclosure)—were collected across 50 plots in April and October, 2023, along with functional traits from 24 species. These traits include root length, leaf area, specific leaf area, clonality, life history, and seed mass. HMSC results revealed that soil properties and precipitation were the primary drivers of community structure, while sand height and elevation had a moderate influence. In contrast, competition and grazing played relatively minor roles. Species responses to environmental covariates were heterogeneous: soil fertility and texture had mixed effects, benefiting some species while limiting others; sand encroachment and precipitation variability generally had negative impacts, whereas grazing exclusion favored many species. A weak phylogenetic signal was recorded, indicating that community assembly was driven more by environmental filtering than by shared evolutionary history. Functional trait responses to environmental variation reflected plant strategies that balanced resource acquisition and conservation. Specifically, seed mass, leaf area, and root length increased under higher soil moisture and nutrient availability but declined in response to salinity, precipitation variability, and sand height. Clonality and perennial life history traits enhanced the survival of plant species under harsh conditions. Overall, this study provides a holistic understanding of community assembly processes in the El Bayadh steppe and offers valuable insights for ecosystem management and restoration in arid and degraded ecosystem environments.
Patrick-Nino OLOUMANE , Carlo PREVIL , Wael El ZEREY , Asma El ZEREY-BELASKRI . Applying joint species distribution modelling to assess the relative influence of ecological filters on community assembly in the El Bayadh steppe, Algeria[J]. Journal of Arid Land, 2025 , 17(7) : 979 -996 . DOI: 10.1007/s40333-025-0082-y
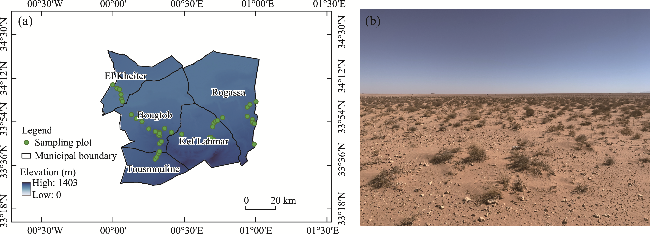
Fig. 1 Geographical distribution of vegetation sampling plots across the study area (a) and example of a degraded steppe in Kef Lahmar municipality, El Bayadh Province, Algeria (b) |
Table 1 Functional traits and their ecological function |
| Functional trait | Ecological function | Ecological filter |
|---|---|---|
| Specific leaf area | Acquisition, retention, and growth | Environmental |
| Leaf area | Acquisition and retention | Environmental |
| Seed mass | Dispersion and germination | Dispersal |
| Life history | Persistence | Biotic |
| Clonality | Space acquisition | Biotic |
| Root length | Resource acquisition | Biotic |
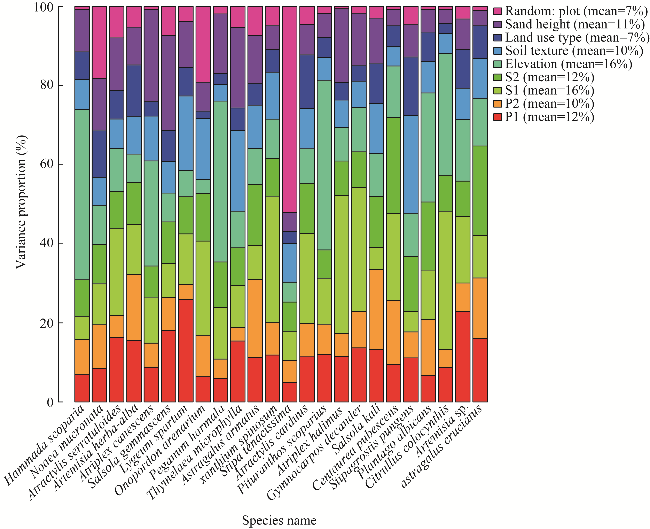
Fig. 2 Variance Partitioning. The values in the legend summarize the mean variance proportion of covariates over the species. Random: plot, random effect at the plot level; P1, precipitation principal component related to north-south humidity gradient; P2, precipitation principal component related to seasonal variability; S1, soil principal component reflecting the gradient of salinity and electrical conductivity; S2, soil principal component reflecting fertility and moisture. |
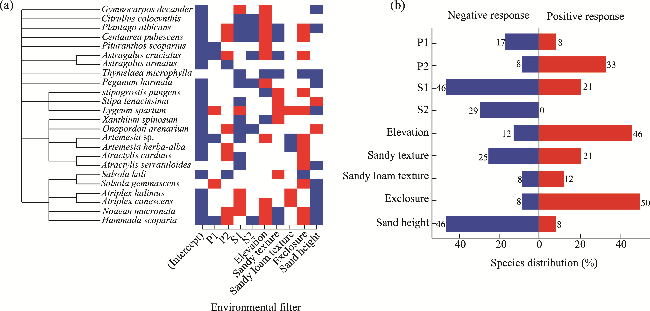
Fig. 3 Species responses to environmental filters. (a), directional trend; (b), response distribution for each environmental filter. The species are ordered according to their phylogeny as illustrated by the phylogenetic tree. The red color indicates a positive response of species with at least a 75% posterior probability, while the blue color indicates a negative response of species with at least a 75% posterior probability. A weak statistical relevance is shown by the white color. |
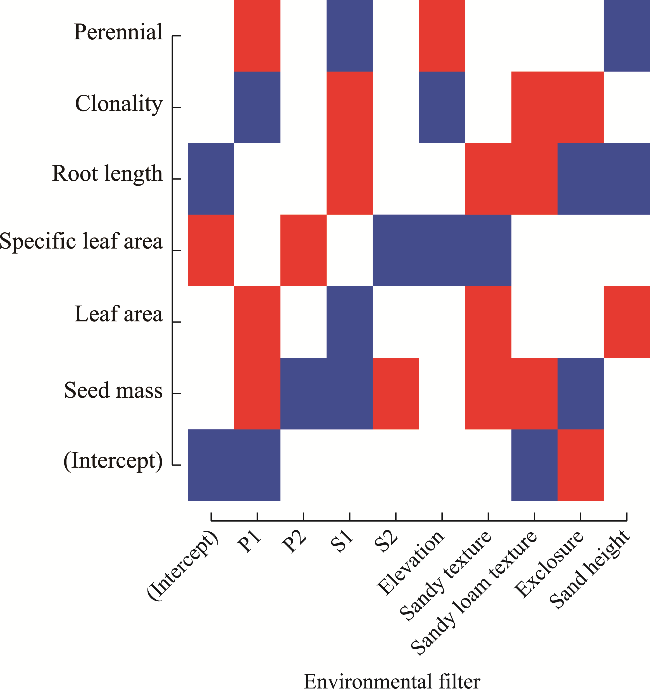
Fig. 4 Responses of functional traits to environmental filters. The intercept represents the expected value of the trait when the environmental variable equals zero. Although the value zero may not always be ecologically meaningful, the intercept is included in the regression model for mathematical completeness and accurate estimation of the relationship. |
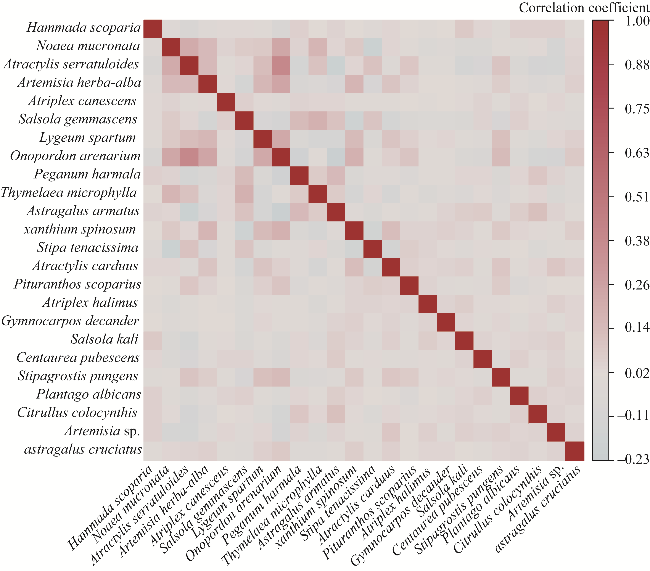
Fig. 5 Species-to-species association matrix highlighting the interspecific relationships |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}