
Climate and topography regulate the spatial pattern of soil salinization and its effects on shrub community structure in Northwest China
Received date: 2024-01-31
Revised date: 2024-05-28
Accepted date: 2024-05-30
Online published: 2025-08-14
DU Lan , TIAN Shengchuan , ZHAO Nan , ZHANG Bin , MU Xiaohan , TANG Lisong , ZHENG Xinjun , LI Yan . [J]. Journal of Arid Land, 2024 , 16(7) : 925 -942 . DOI: 10.1007/s40333-024-0060-9
Soil salinization may affect biodiversity and species composition, leading to changes in the plant community structure. However, few studies have explored the spatial pattern of soil salinization and its effects on shrub community structure at the ecosystem scale. Therefore, we conducted a transect sampling of desert shrublands in Northwest China during the growing season (June-September) in 2021. Soil salinization (both the degree and type), shrub community structure (e.g., shrub density and height), and biodiversity parameters (e.g., Simpson diversity, Margalf abundance, Shannon-Wiener diversity, and Pielou evenness indices) were used to assess the effects of soil salinization on shrub community structure. The results showed that the primary degree of soil salinization in the study area was light salinization, with the area proportion of 69.8%. Whereas the main type of soil salinization was characterized as sulfate saline soil, also accounting for 69.8% of the total area. Notably, there was a significant reduction in the degree of soil salinization and a shift in the type of soil salinization from chloride saline soil to sulfate saline soil, with an increase in longitude. Regional mean annual precipitation (MAP), mean annual evapotranspiration (MAE), elevation, and slope significantly contributed to soil salinization and its geochemical differentiation. As soil salinization intensified, shrub community structure displayed increased diversity and evenness, as indicated by the increases in the Simpson diversity, Shannon-Wiener diversity, and Pielou evenness indices. Moreover, the succulent stems and leaves of Chenopodiaceae and Tamaricaceae exhibited clear advantages under these conditions. Furthermore, regional climate and topography, such as MAP, MAE, and elevation, had greater effects on the distribution of shrub plants than soil salinization. These results provide a reference for the origin and pattern of soil salinization in drylands and their effects on the community structure of halophyte shrub species.
Key words: soil salinization; halophytes; shrubland; climate change; biodiversity; drylands; Northwest China
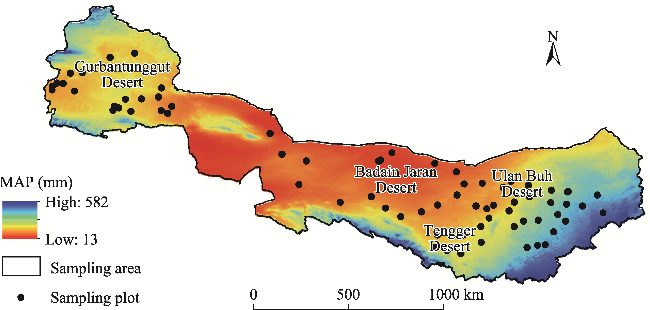
Fig. 1 Geographical overview of the study area and location of the sampling plots. MAP, mean annual precipitation. Note that the figure is based on the standard map (GS(2023)2767) from the Standard Map Service System (http://bzdt.ch.mnr.gov.cn/index.html) marked by the Ministry of Natural Resources of the People's Republic of China, and the standard map has not been modified. |
Table S1 Summary of the shrub plant communities in the 72 sampling plots in Northwest China |
| Plot ID | Latitude | Longitude | Elevation (m) | Dominant species |
|---|---|---|---|---|
| A | 45°21′20′′N | 85°00′17′′E | 280 | Nitraria tangutorum, Tamarix ramosissima, Anabasis brevifolia, and Haloxylon ammodendron# |
| B | 45°10′30′′N | 84°58′58′′E | 290 | Nitraria tangutorum, Tamarix ramosissima, Reaumuria songarica, and Haloxylon ammodendron# |
| C | 45°29′50′′N | 85°30′24′′E | 260 | Nitraria tangutorum, Tamarix ramosissima, Anabasis brevifolia, Reaumuria songarica, and Haloxylon ammodendron# |
| D | 45°30′20′′N | 85°12′27′′E | 280 | Anabasis brevifolia, Haloxylon ammodendron#, and Kalidium foliatum |
| E | 45°07′22′′N | 86°01′41′′E | 340 | Tamarix ramosissima#, Reaumuria songarica, Nitraria tangutorum, and Haloxylon ammodendron |
| F | 44°09′48′′N | 88°42′32′′E | 610 | Tamarix ramosissima#, Anabasis brevifolia, Reaumuria songarica, Haloxylon ammodendron, Suaeda microphylla, and Kalidium foliatum |
| G | 44°45′42′′N | 89°13′05′′E | 540 | Nitraria tangutorum, Anabasis brevifolia, Ephedra major, Reaumuria songarica, Calligonum mongolicum, and Haloxylon ammodendron# |
| H | 44°51′08′′N | 90°01′06′′E | 650 | Haloxylon ammodendron# and Halostachys caspica |
| I | 44°23′45′′N | 90°38′47′′ | 830 | Anabasis brevifolia, Reaumuria songarica, Haloxylon ammodendron#, and Kalidium foliatum |
| J | 44°04′28′′N | 90°26′52′′E | 920 | Nitraria tangutorum# and Ceratoides latens |
| K | 44°12′01′′N | 90°08′37′′E | 760 | Haloxylon persicum#, Atraphaxis bracteata, Anabasis brevifolia, Calligonum mongolicum, Artemisia ordosica, Haloxylon ammodendron, and Ceratoides latens |
| L | 45°17′14′′N | 90°09′54′′E | 1200 | Anabasis brevifolia, Reaumuria songarica, and Haloxylon ammodendron# |
| M | 46°55′54′′N | 88°54′11′′E | 830 | Anabasis brevifolia and Haloxylon ammodendron# |
| N | 46°43′28′′N | 87°43′32′′E | 570 | Salsola laricifolia, Tamarix ramosissima, Halimodendron halodendron, Reaumuria songarica, Haloxylon ammodendron#, and Ceratoides latens |
| O | 45°58′52′′N | 85°50′28′′E | 280 | Tamarix ramosissima#, Anabasis brevifolia, Reaumuria songarica, and Haloxylon ammodendron |
| P | 46°00′43′′N | 86°24′44′′E | 320 | Tamarix ramosissima, Nitraria roborowskii, Nitraria sibirica, Haloxylon ammodendron#, and Kalidium foliatum |
| Q | 41°49′04′′N | 97°02′10′′E | 1740 | Nitraria tangutorum, Reaumuria songarica, Haloxylon ammodendron#, and Halostachys caspica |
| R | 42°12′21′′N | 101°05′60′′E | 920 | Tamarix ramosissima# |
| S | 41°52′03′′N | 100°32′52′′E | 970 | Nitraria tangutorum, Tamarix ramosissima#, and Lycium ruthenicum |
| T | 41°48′56′′N | 100°28′05′′E | 980 | Nitraria tangutorum, Tamarix ramosissima#, Reaumuria songarica, and Calligonum mongolicum |
| U | 41°41′49′′N | 103°07′57′′E | 1010 | Reaumuria songarica# |
| V | 41°17′58′′N | 104°08′37′′E | 800 | Nitraria tangutorum, Tamarix ramosissima#, Haloxylon ammodendron, and Kalidium foliatum |
| W | 40°43′59′′N | 104°30′36′′E | 1320 | Nitraria tangutorum# and Reaumuria songarica |
| X | 40°51′29′′N | 106°44′31′′E | 1040 | Haloxylon ammodendron# |
| Y | 40°33′55′′N | 106°22′33′′E | 1040 | Nitraria tangutorum, Tamarix ramosissima, and Haloxylon ammodendron# |
| Z | 40°34′24′′N | 106°22′00′′E | 1050 | Nitraria tangutorum, Sarcozygium xanthoxylum#, and Ammopiptanthus mongolicus |
| A1 | 40°45′41′′N | 105°22′37′′E | 1180 | Nitraria tangutorum, Potaninia mongolica, Reaumuria songarica, and Kalidium foliatum# |
| B1 | 40°39′50′′N | 107°33′49′′E | 1040 | Artemisia ordosica and Salix cheilophila# |
| C1 | 40°25′48′′N | 108°39′24′′E | 1120 | Artemisia ordosica and Salix cheilophila# |
| D1 | 40°21′48′′N | 109°25′23′′E | 1080 | Hedysarum scoparium#, Caragana sinica, and Artemisia ordosica |
| E1 | 40°22′20′′N | 109°26′35′′E | 1070 | Caragana sinica, Artemisia ordosica, and Salix cheilophila# |
| F1 | 40°12′20′′N | 110°50′23′′E | 1040 | Caragana sinica, Artemisia ordosica, and Salix cheilophila# |
| Plot ID | Latitude | Longitude | Elevation (m) | Dominant species |
| G1 | 39°21′48′′N | 111°06′56′′E | 890 | Caragana sinica# and Artemisia ordosica |
| H1 | 39°42′59′′N | 105°54′31′′E | 1170 | Caragana sinica# and Artemisia ordosica |
| I1 | 39°41′21′′N | 110°08′16′′E | 1490 | Caragana sinica, Hippophae rhamnoides#, and Kalidium foliatum |
| J1 | 39°46′17′′N | 109°22′26′′E | 1370 | Caragana sinica, Artemisia ordosica, and Salix cheilophila# |
| K1 | 39°50′28′′N | 108°37′35′′E | 1370 | Atraphaxis bracteata, Hedysarum scoparium#, Caragana sinica, and Artemisia ordosica |
| L1 | 40°07′37′′N | 107°36′50′′E | 1200 | Caragana sinica# and Ceratoides latens |
| M1 | 40°15′23′′N | 107°04′14′′E | 1140 | Nitraria tangutorum, Sarcozygium xanthoxylum, Caragana sinica#, Ammopiptanthus mongolicus, Artemisia ordosica, Tetraena mongolica, Reaumuria songarica, Sabina vulgaris, and Reaumuria songarica |
| N1 | 39°52′26′′N | 106°52′57′′E | 1230 | Sarcozygium xanthoxylum, Ammopiptanthus mongolicus, Artemisia ordosica, Tetraena mongolica, Potaninia mongolica, and Sabina vulgaris# |
| O1 | 38°58′56′′N | 107°19′33′′E | 1180 | Nitraria tangutorum, Reaumuria songarica, Ammopiptanthus mongolicus#, Kalidium foliatum, Reaumuria songarica, and Kochia prostrata |
| P1 | 38°42′47′′N | 106°53′08′′E | 1240 | Artemisia ordosica# |
| Q1 | 37°43′37′′N | 107°29′50′′E | 1330 | Caragana sinica# |
| R1 | 37°49′04′′N | 108°00′03′′E | 1360 | Hedysarum scoparium, Caragana sinica, Artemisia ordosica, Salix cheilophila#, and Amorpha fruticosa |
| S1 | 37°51′12′′N | 108°22′25′′E | 1310 | Caragana sinica, Artemisia ordosica, and Salix cheilophila# |
| T1 | 38°28′02′′N | 108°46′14′′E | 1360 | Caragana sinica# and Artemisia ordosica |
| U1 | 38°58′13′′N | 109°19′11′′E | 1310 | Sabina vulgaris#, Caragana sinica, Artemisia ordosica, and Salix cheilophila |
| V1 | 39°16′32′′N | 108°57′02′′E | 1310 | Hedysarum scoparium, Caragana sinica, Artemisia ordosica, Hippophae rhamnoides, and Salix cheilophila# |
| W1 | 39°01′04′′N | 108°01′33′′E | 1370 | Artemisia ordosica# |
| X1 | 39°27′27′′N | 106°38′37′′E | 1230 | Nitraria tangutorum, Sarcozygium xanthoxylum, Caragana sinica#, Potaninia mongolica, Reaumuria songarica, Ammopiptanthus mongolicus, Tetraena mongolica, and Ceratoides latens |
| Y1 | 39°06′27′′N | 105°42′06′′E | 1660 | Hedysarum scoparium#, Calligonum mongolicum, and Artemisia ordosica |
| Z1 | 37°58′35′′N | 105°20′22′′E | 1330 | Nitraria tangutorum# and Reaumuria songarica |
| A2 | 37°26′19′′N | 104°22′04′′E | 1640 | Nitraria tangutorum, Reaumuria songarica#, and Salsola passerina |
| B2 | 37°34′29′′N | 103°42′45′′E | 1750 | Nitraria tangutorum#, Caragana sinica, and Artemisia ordosica |
| C2 | 37°43′47′′N | 103°08′52′′E | 1660 | Hedysarum scoparium# and Artemisia ordosica |
| D2 | 38°18′43′′N | 103°15′36′′E | 1470 | Nitraria tangutorum# and Reaumuria songarica |
| E2 | 38°16′43′′N | 103°55′36′′E | 1400 | Nitraria tangutorum# |
| F2 | 38°18′06′′N | 104°28′40′′E | 1360 | Nitraria tangutorum# and Kalidium foliatum |
| G2 | 38°43′12′′N | 105°18′17′′E | 1280 | Hedysarum scoparium# and Artemisia ordosica |
| H2 | 39°33′03′′N | 105°35′52′′E | 1050 | Nitraria tangutorum#, Ammopiptanthus mongolicus, Calligonum mongolicum, and Artemisia ordosica |
| I2 | 39°41′07′′N | 105°04′52′′E | 1320 | Nitraria tangutorum, Reaumuria songarica, Kalidium foliatum, Salsola laricifolia#, and Salsola passerina |
| J2 | 40°12′54′′N | 104°10′50′′E | 1440 | Nitraria tangutorum# and Reaumuria songarica |
| K2 | 39°44′01′′N | 103°15′37′′E | 1280 | Nitraria tangutorum and Artemisia ordosica# |
| L2 | 39°24′56′′N | 102°28′47′′E | 1340 | Nitraria tangutorum, Sarcozygium xanthoxylum#, and Artemisia ordosica |
| M2 | 39°11′39′′N | 101°30′39′′E | 1420 | Nitraria tangutorum, Reaumuria songarica, Kalidium foliatum#, and Salsola passerina |
| N2 | 39°35′19′′N | 100°47′49′′E | 1510 | Nitraria tangutorum, Reaumuria songarica, Kalidium foliatum#, and Salsola passerina |
| Plot ID | Latitude | Longitude | Elevation (m) | Dominant species |
| O2 | 40°08′19′′N | 100°06′41′′E | 1240 | Nitraria tangutorum#, Reaumuria songarica, and Calligonum mongolicum |
| P2 | 39°51'54''N | 98°38'58''E | 1400 | Nitraria tangutorum, Reaumuria songarica#, and Calligonum mongolicum |
| Q2 | 40°43′05′′N | 96°40′42′′E | 1510 | Reaumuria songarica# |
| R2 | 42°08′43′′N | 95°52′39′′E | 1820 | Reaumuria songarica and Haloxylon ammodendron# |
| S2 | 43°7′13′′N | 95°18′16′′E | 1130 | Ephedra major# |
| T2 | 44°12′53′′N | 87°51′39′′E | 490 | Sarcozygium xanthoxylum, Tamarix ramosissima#, Reaumuria songarica, and Kalidium foliatum |
| U2 | 44°20′42′′N | 88°08′06′′E | 480 | Tamarix ramosissima#, Anabasis brevifolia, Reaumuria songarica, and Haloxylon ammodendron |
| V2 | 44°43′60′′N | 88°27′00′′E | 4665 | Nitraria roborowskii, Tamarix ramosissima#, Reaumuria songarica, Kalidium foliatum, Halimodendron halodendron, and Nitraria sibirica |
| W2 | 44°23′52′′N | 87°55′34′′E | 451 | Haloxylon persicum, Tamarix ramosissima, and Haloxylon ammodendron# |
| X2 | 44°22′45′′N | 87°56′38′′E | 470 | Tamarix ramosissima#, Reaumuria songarica, and Haloxylon ammodendron |
| Y2 | 44°24′56′′N | 87°54′58′′E | 454 | Haloxylon persicum#, Reaumuria songarica, and Haloxylon ammodendron |
| Z2 | 44°22′22′′N | 87°56′15′′E | 460 | Haloxylon persicum and Haloxylon ammodendron# |
Note: #, constructive species in each sampling plot. The plot ID numbers are the same in Table S2. |
Table 1 Classification of soil salinization |
| Classification system | Soil type | Criteria |
|---|---|---|
| Classification system based on American Soil Salinity Classification System# | Non-salinized soil | TS≤5 mg/kg |
| Slightly salinized soil | 5 mg/kg<TS≤10 mg/kg | |
| Moderately salinized soil | 10 mg/kg<TS≤15 mg/kg | |
| Severely salinized soil | 15 mg/kg<TS≤20 mg/kg | |
| Classification system based on anion of salt types## | Sulfate saline soil | Cl-/SO42-≤0.2 |
| Chloride-sulfate saline soil | 0.2<Cl-/SO42-≤1.0 | |
| Sulfate-chloride saline | 1.0<Cl-/SO42-≤4.0 | |
| Chloride saline soil | Cl-/SO42->4.0 |
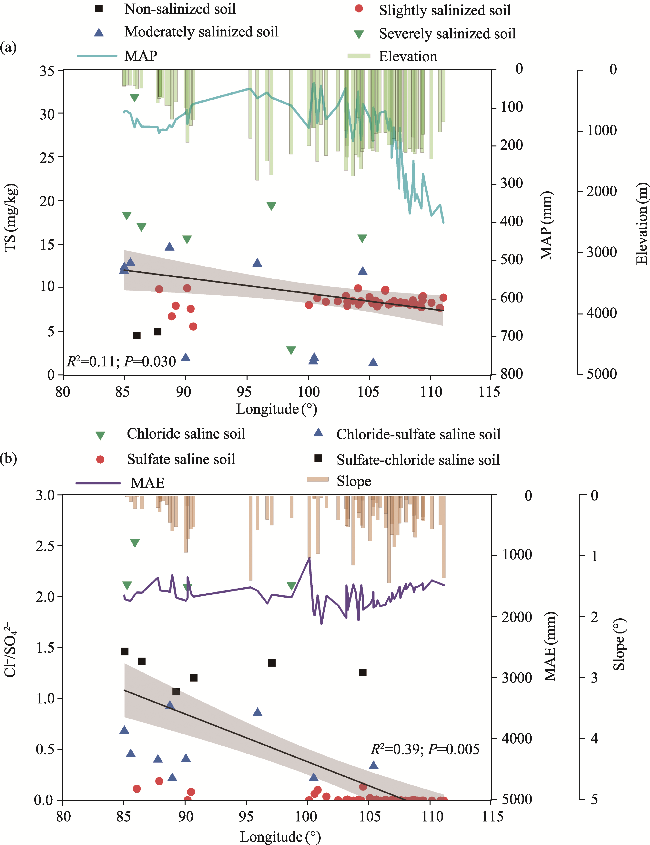
Fig. 2 Variations of soil salinization degree, MAP, and elevation (a) and soil salinization type, mean annual evapotranspiration (MAE), and slope (b) along the longitudinal gradient. Note that soil salinization degree and type are expressed as soil total salt (TS) and Cl-/SO42-, respectively. Black solid line indicates significant relationship between soil salinization and longitude (P<0.050). Shaded area represents 95% confidence interval. R2 and P values represent the linear relationship between soil salinization and longitude. |
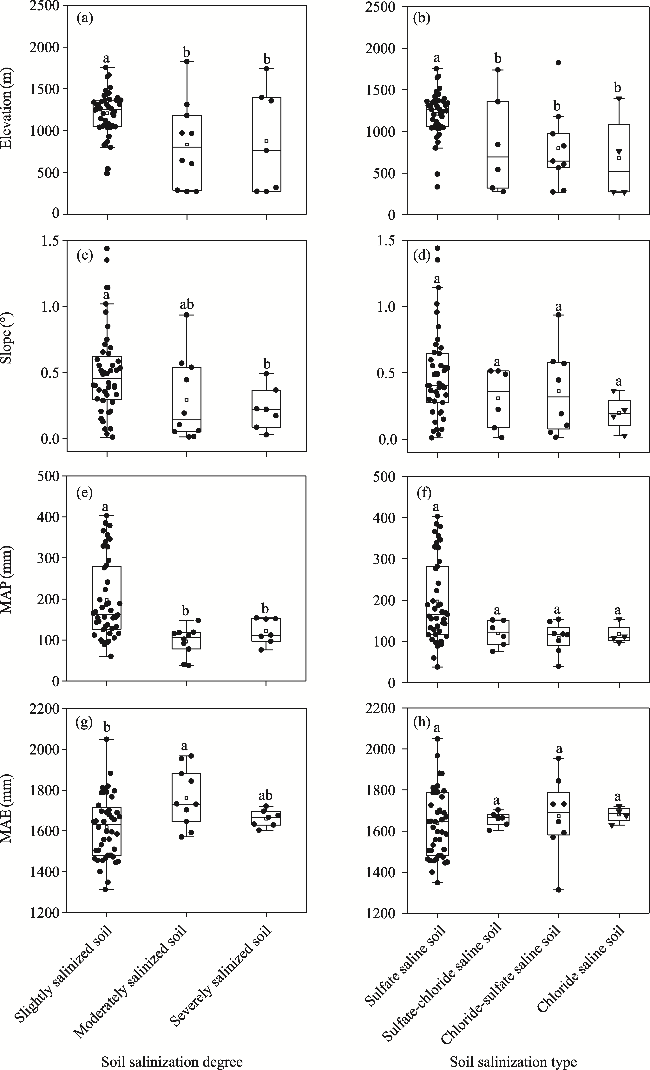
Fig. 3 Effects of topographical (a, b, c, and d) and climatic (e, f, g, and h) factors on soil salinization degree and type. Different lowercase letters indicate significant differences at the P<0.050 level. Since there were few sampling plots with non-salinized soil (n=2, where n is the total number of sampling plots), they were not plotted in this figure. Black dots indicate the data points of elevation, slope, MAP, and MAE. Box boundaries indicate the 25th and 75th percentiles, and whiskers below and above the box indicate the 10th and 90th percentiles, respectively. The black horizontal line within each box indicates the median of data points. |
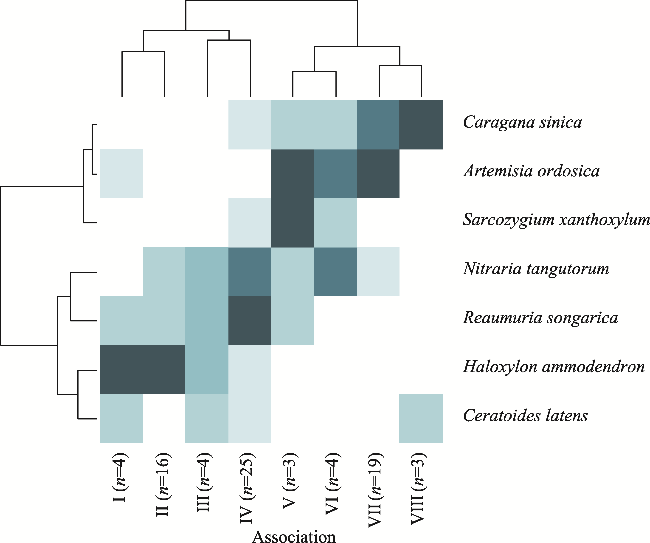
Fig. 4 TWINSPAN tree classification of the 78 sampling plots. The heat map shows data on the abundance of the dominant species in the corresponding quadrats below the cluster tree. The clustering results of the quadrats (four-layer classification) are at the top of the figure, and the clustering results of shrub species are on the left of the figure. n is the total number of sampling plots. The dominant species and sampling plots in each association are shown in Table S2. |
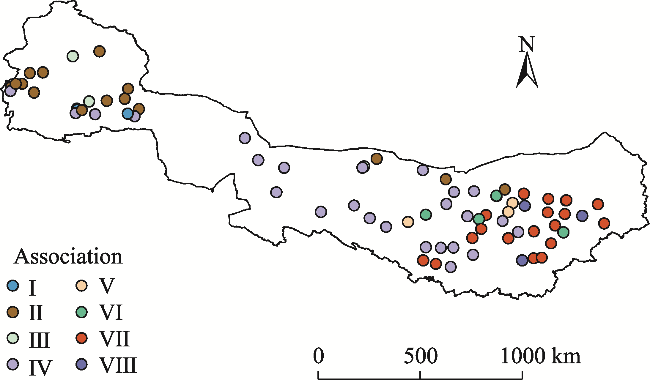
Fig. S1 Geographical distribution of shrub associations along the longitudinal gradient in Northwest China |
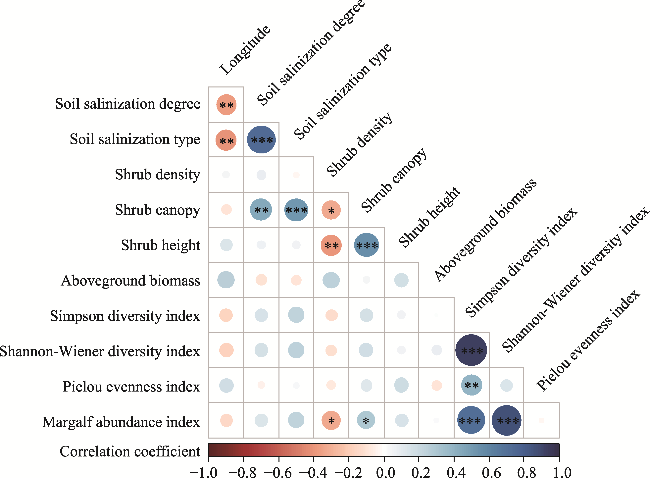
Fig. S2 Pearson's correlations among longitude, soil salinization, and parameters of shrub community structure. The size of the colored circle indicates the degree of correlation. *, P<0.050 level; **, P<0.010 level; ***, P<0.001 level. |
Table S2 Dominant species and sampling plots in each association |
| Association | Cluster name | Sampling plots |
|---|---|---|
| I | Haloxylon ammodendron+Haloxylon persicum+Ceratoides latens | K, W2, Y2, and Z2 |
| II | Haloxylon ammodendron+Anabasis brevifolia+Tamarix ramosissima | A, C, D, E, G, H, I, L, M, O, P, R, U2, V, X, and X2 |
| III | Nitraria tangutorum+Reaumuria songarica+ Tamarix ramosissima | N, S, V2, and Y |
| IV | Reaumuria songarica+Nitraria tangutorum+Salsola passerina | A1, A2, B, D2, E2, F, F2, I2, J, J2, M2, N2, O1, O2, P2, Q, Q2, R2, S2, T, T2, U, W, X1, and Z1 |
| V | Reaumuria songarica+Nitraria tangutorum+Kalidium foliatum | L2, M1, and N1 |
| VI | Calligonum mongolicum+Reaumuria songarica+Halostachys caspica | H2, K2, U1, and Z |
| VII | Reaumuria songarica+Tetraena mongolica+ Salix cheilophila | B1, B2, C1, C2, D1, E1, F1, G1, G2, H1, J1, K1, P1, R1, S1, T1, V1, W1, and Y1 |
| VIII | Tetraena mongolica+Suaeda microphylla | I1, L1, and Q1 |
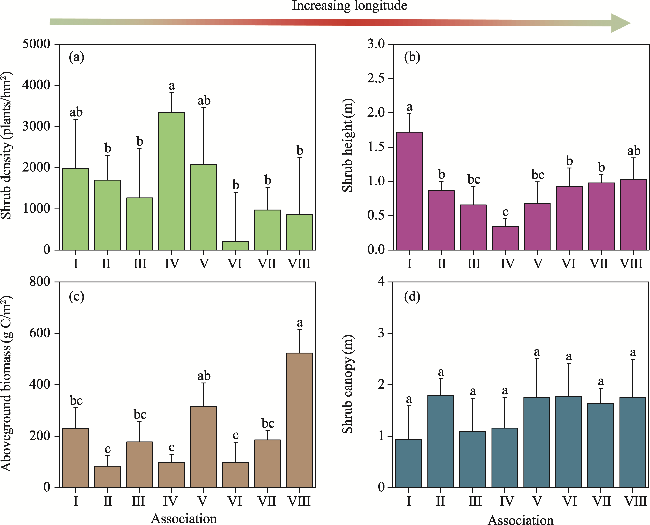
Fig. 5 Comparison of shrub density (a), shrub height (b), aboveground biomass (c), and shrub canopy (d) among the eight shrub associations. Different lowercase letters indicate significant differences among different associations at the P<0.050 level. Bars mean standard errors. |
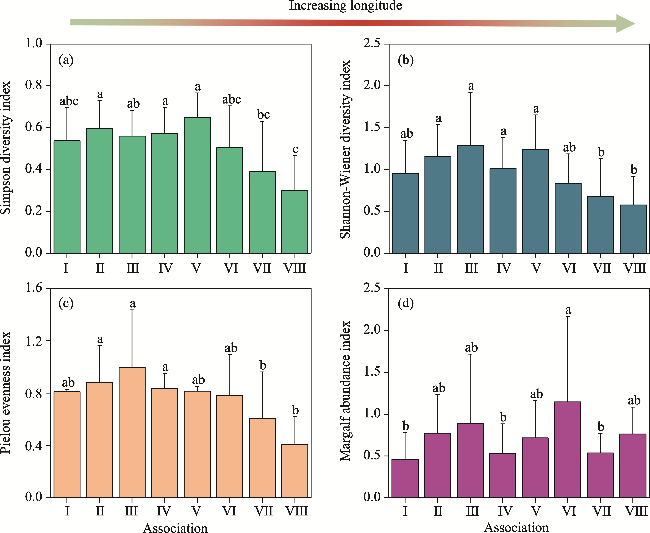
Fig. 6 Comparison of the Simpson diversity (a), Shannon-Wiener diversity (b), Pielou evenness (c), and Margalf abundance (d) indices among the eight shrub associations. Different lowercase letters indicate significant differences among different associations at the P<0.050 level. Bars mean standard errors. |
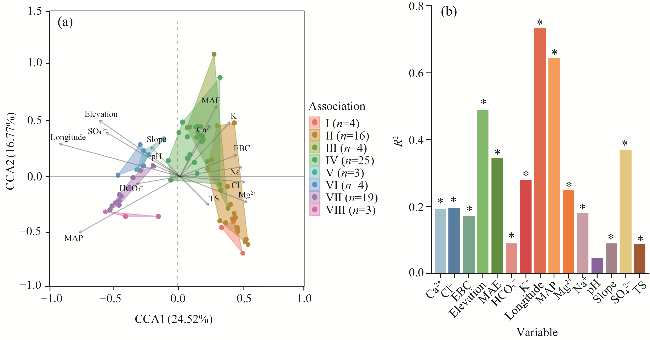
Fig. 7 Canonical correlation analysis (CCA) of the eight shrub associations (a) and their affecting variables of soil salinization, climate, and topography (b). In the left panel, different colors and areas correspond to the distribution ranges of shrub associations. CCA1 and CCA2 are the first and second axes of CCA, respectively. EBC, exchangeable base cation. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}