
Historical tillage promotes grass-legume mixtures establishment and accelerates soil microbial activity and organic carbon decomposition
The first and second authors contributed equally to this work.
Received date: 2024-03-06
Revised date: 2024-06-03
Accepted date: 2024-06-18
Online published: 2025-08-14
ZHOU Jiqiong , GONG Jinchao , WANG Pengsen , SU Yingying , LI Xuxu , LI Xiangjun , LIU Lin , BAI Yanfu , MA Congyu , WANG Wen , HUANG Ting , YAN Yanhong , ZHANG Xinquan . [J]. Journal of Arid Land, 2024 , 16(7) : 910 -924 . DOI: 10.1007/s40333-024-0021-3
Perennial grass-legume mixtures have been extensively used to restore degraded grasslands, increasing grassland productivity and forage quality. Tillage is crucial for seedbed preparation and sustainable weed management for the establishment of grass-legume mixtures. However, a common concern is that intensive tillage may alter soil characteristics, leading to losses in soil organic carbon (SOC). We investigated the plant community composition, SOC, soil microbial biomass carbon (MBC), soil enzyme activities, and soil properties in long-term perennial grass-legume mixtures under two different tillage intensities (once and twice) as well as in a fenced grassland (FG). The establishment of grass-legume mixtures increased plant species diversity and plant community coverage, compared with FG. Compared with once tilled grassland (OTG), twice tilled grassland (TTG) enhanced the coverage of high-quality leguminous forage species by 380.3%. Grass-legume mixtures with historical tillage decreased SOC and dissolved organic carbon (DOC) concentrations, whereas soil MBC concentrations in OTG and TTG increased by 16.0% and 16.4%, respectively, compared with FG. TTG significantly decreased the activity of N-acetyl-β-D-glucosaminidase (NAG) by 72.3%, whereas soil enzyme β-glucosidase (βG) in OTG and TTG increased by 55.9% and 27.3%, respectively, compared with FG. Correlation analysis indicated a close association of the increase in MBC and βG activities with the rapid decline in SOC. This result suggested that MBC was a key driving factor in soil carbon storage dynamics, potentially accelerating soil carbon cycling and facilitating biogeochemical cycling. The establishment of grass-legume mixtures effectively improves forage quality and boosts plant diversity, thereby facilitating the restoration of degraded grasslands. Although tillage assists in establishing legume-grass mixtures by controlling weeds, it accelerates microbial activity and organic carbon decomposition. Our findings provide a foundation for understanding the process and effectiveness of restoration management in degraded grasslands.
Table 1 Plant species of fenced grassland (FG), once tilled grassland (OTG), and twice tilled grassland (TTG) |
| Type of grassland | Plant species | Family | Relative abundance (%) |
|---|---|---|---|
| FG | Imperata cylindrica (L.) P. Beauv. | Gramineae | 73.59±4.42 |
| Eragrostis ferruginea (Thunb.) P. Beauv. | Gramineae | 2.56±0.00 | |
| Artemisia dubia Wall. ex Besser subf. intermedia Pamp. | Compositae | 8.21±2.10 | |
| Galium aparine L. | Rubiaceae | 7.80±2.20 | |
| Aster indicus L. | Compositae | 1.97±1.33 | |
| Duchesnea indica (Andrews) Focke | Rosaceae | 2.83±0.00 | |
| Elsholtzia ciliata (Thunb.) Hyl. | Lamiaceae | 4.64±0.00 | |
| Lonicera japonica Thunb. | Caprifoliaceae | 1.11±0.00 | |
| Potentilla chinensis Ser. | Rosaceae | 1.15±0.00 | |
| Origanum vulgare L. | Lamiaceae | 5.00±0.00 | |
| Dichondra micrantha Urb. | Convolvulaceae | 1.23±0.00 | |
| Dactylis glomerata L. | Gramineae | 1.11±0.00 | |
| Trifolium repens L. | Leguminosae | 1.23±0.00 | |
| OTG | Dactylis glomerata L. | Gramineae | 44.60±9.62 |
| Trifolium repens L. | Leguminosae | 6.58±3.00 | |
| Lolium perenne L. | Gramineae | 3.59±0.62 | |
| Cynodon dactylon (L.) Persoon | Gramineae | 6.30±6.10 | |
| Vicia Sativa L. | Leguminosae | 0.46±0.24 | |
| Artemisia dubia Wall. ex Besser subf. intermedia Pamp. | Compositae | 12.09±1.69 | |
| Lonicera japonica Thunb. | Caprifoliaceae | 2.25±1.21 | |
| Dichondra micrantha Urb. | Convolvulaceae | 9.97±7.93 | |
| Eleusine indica (L.) Gaertn. | Gramineae | 5.74±0.00 | |
| Clinopodium chinense (Benth.) Kuntze | Lamiaceae | 2.87±0.00 | |
| Bidens pilosa L. | Compositae | 1.26±0.00 | |
| Hypochaeris ciliata (Thunb.) Makino | Compositae | 9.02±0.00 | |
| Plantago asiatica L. | Plantaginaceae | 1.23±0.00 | |
| Aster indicus L. | Compositae | 0.92±0.00 | |
| TTG | Dactylis glomerata L. | Gramineae | 37.45±7.08 |
| Trifolium repens L. | Leguminosae | 28.45±8.80 | |
| Lolium perenne L. | Gramineae | 2.32±0.00 | |
| Cynodon dactylon (L.) Persoon | Gramineae | 4.62±2.76 | |
| Vicia Sativa L. | Leguminosae | 5.35±0.39 | |
| Sporobolus fertilis (Steud.) Clayton | Gramineae | 4.39±0.00 | |
| Imperata cylindrica (L.) P. Beauv. | Gramineae | 7.71±2.51 | |
| Artemisia dubia Wall. ex Besser subf. intermedia Pamp. | Compositae | 5.30±2.66 | |
| Lonicera japonica Thunb. | Caprifoliaceae | 3.97±0.00 | |
| Dichondra micrantha Urb. | Convolvulaceae | 1.59±0.09 | |
| Hypochaeris ciliata (Thunb.) Makino | Compositae | 1.15±0.00 | |
| Rumex acetosa L. | Polygonaceae | 3.84±2.14 | |
| Aster indicus L. | Compositae | 1.15±0.00 | |
| Potentilla chinensis Ser. | Rosaceae | 1.72±0.00 | |
| Elsholtzia ciliata (Thunb.) Hyl. | Lamiaceae | 1.75±0.00 | |
| Artemisia argyi H. Lév. & Vaniot | Compositae | 0.62±0.00 |
Note: Mean±SE. |
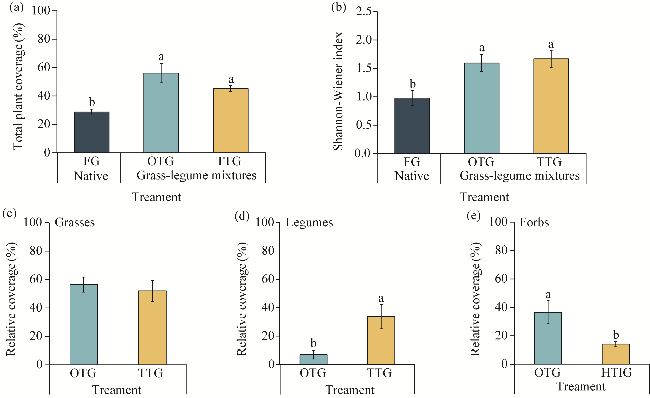
Fig. 1 Total plant coverage (a), Shannon-Wiener index (b), and relative coverages of grasses (c), legumes (d), and forbs (e) under different grassland treatments. FG, fenced grassland; OTG, once tilled grassland; TTG, twice tilled grassland. Different lowercase letters indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. The abbreviations are the same as in the following figures. |
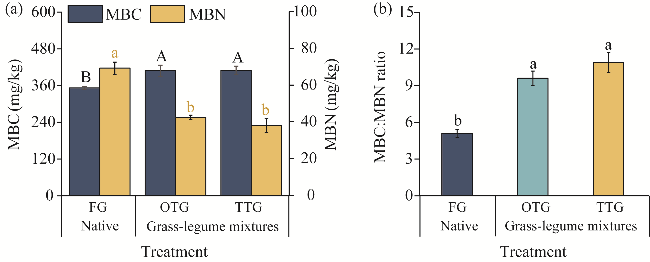
Fig. 2 Concentrations of MBC and MBN (a) and MBC:MBN ratio (b) under different grassland treatments. MBC, microbial biomass carbon; MBN, microbial biomass nitrogen. Different uppercase or lowercase letters within the same parameter indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. |
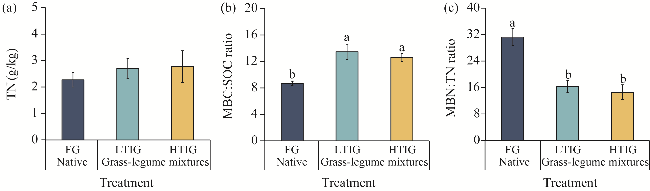
Fig. 3 Concentration of TN (a), MBC:SOC ratio (b), and MBN:TN ratio (c) under different grassland treatments. TN, total nitrogen; SOC, soil organic carbon. Different lowercase letters indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. |
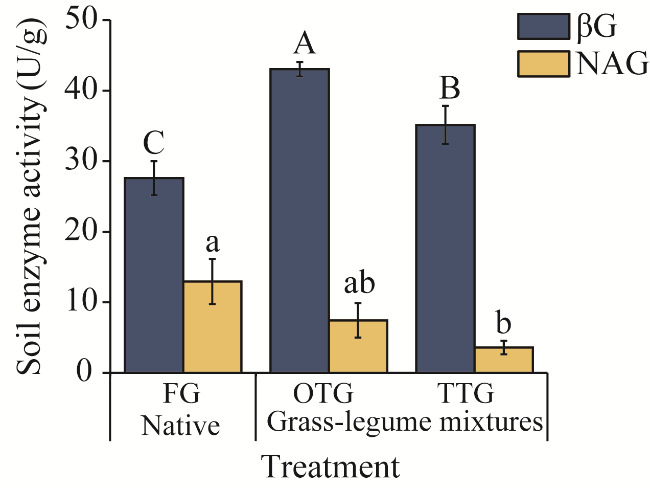
Fig. 4 Variation in soil enzyme activity under different grassland treatments. NAG, N-acetyl-β-D-glucosaminidase; βG, β-glucosidase. Different uppercase or lowercase letters within the same parameter indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. |
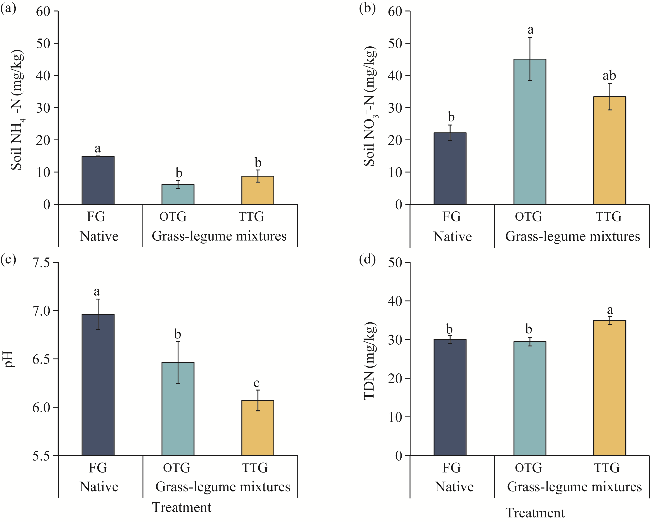
Fig. 5 Concentrations of soil ammonium (NH4+-N; a), nitrate (NO3--N; b), pH (c), and TDN (d) under different grassland treatments. TDN, total dissolved nitrogen. Different lowercase letters indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. |
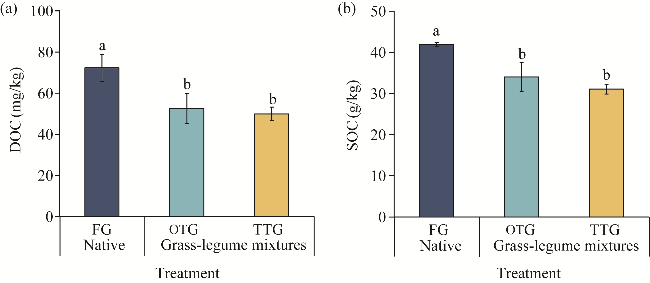
Fig. 6 Concentrations of DOC (a) and SOC (b) under different grassland treatments. DOC, dissolved organic carbon. Different lowercase letters indicate significant differences among different treatments at P<0.05 level. Bars are standard errors. |
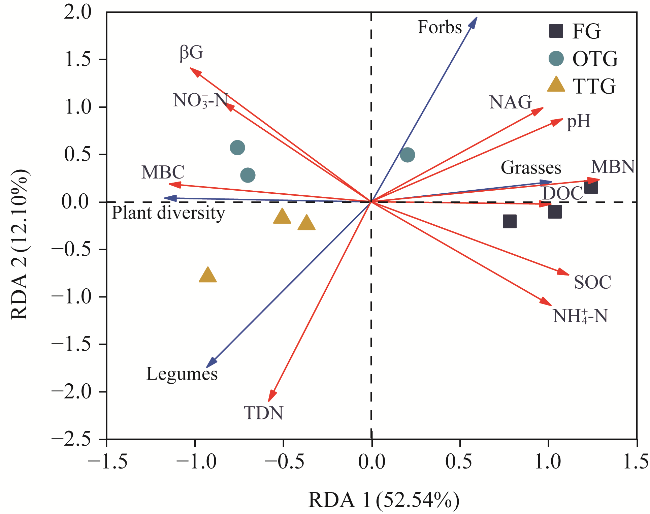
Fig. 7 Redundancy analysis (RDA) of plant composition, soil microbial biomass, soil properties, and soil enzyme activity. Red arrows indicate soil factors, and blue arrows indicate plant characteristics. |
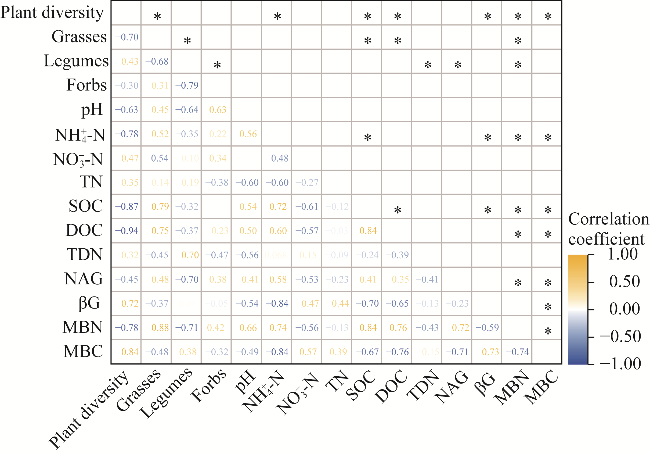
Fig. 8 Heatmap of the correlation of plant community, soil microbial biomass, soil property, and enzyme activity. *, P<0.05 level. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
NBSC (National Bureau of Statistics of China). 2009. Input-Output Tables of China 2007. Beijing: National Bureau of Statistics of China. (in Chinese)
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}