
Impacts of extreme climate and vegetation phenology on net primary productivity across the Qinghai- Xizang Plateau, China from 1982 to 2020
Received date: 2024-07-15
Revised date: 2024-12-21
Accepted date: 2024-12-25
Online published: 2025-08-13
SUN Huaizhang , ZHAO Xueqiang , CHEN Yangbo , LIU Jun . [J]. Journal of Arid Land, 2025 , 17(3) : 350 -367 . DOI: 10.1007/s40333-025-0075-x
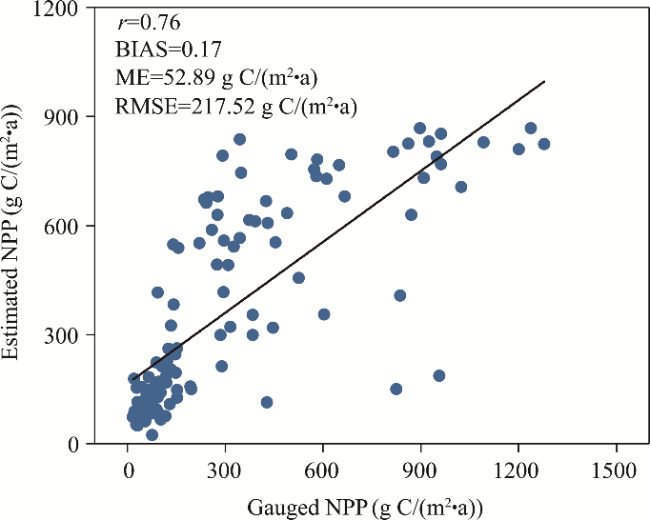
The net primary productivity (NPP) is an important indicator for assessing the carbon sequestration capacities of different ecosystems and plays a crucial role in the global biosphere carbon cycle. However, in the context of the increasing frequency, intensity, and duration of global extreme climate events, the impacts of extreme climate and vegetation phenology on NPP are still unclear, especially on the Qinghai-Xizang Plateau (QXP), China. In this study, we used a new data fusion method based on the MOD13A2 normalized difference vegetation index (NDVI) and the Global Inventory Modeling and Mapping Studies (GIMMS) NDVI3g datasets to obtain a NDVI dataset (1982-2020) on the QXP. Then, we developed a NPP dataset across the QXP using the Carnegie-Ames-Stanford Approach (CASA) model and validated its applicability based on gauged NPP data. Subsequently, we calculated 18 extreme climate indices based on the CN05.1 dataset, and extracted the length of vegetation growing season using the threshold method and double logistic model based on the annual NDVI time series. Finally, we explored the spatiotemporal patterns of NPP on the QXP and the impact mechanisms of extreme climate and the length of vegetation growing season on NPP. The results indicated that the estimated NPP exhibited good applicability. Specifically, the correlation coefficient, relative bias, mean error, and root mean square error between the estimated NPP and gauged NPP were 0.76, 0.17, 52.89 g C/(m2•a), and 217.52 g C/(m2•a), respectively. The NPP of alpine meadow, alpine steppe, forest, and main ecosystem on the QXP mainly exhibited an increasing trend during 1982-2020, with rates of 0.35, 0.38, 1.40, and 0.48 g C/(m2•a), respectively. Spatially, the NPP gradually decreased from southeast to northwest across the QXP. Extreme climate had greater impact on NPP than the length of vegetation growing season on the QXP. Specifically, the increase in extremely-wet-day precipitation (R99p), simple daily intensity index (SDII), and hottest day (TXx) increased the NPP in different ecosystems across the QXP, while the increases in the cold spell duration index (CSDI) and warm spell duration index (WSDI) decreased the NPP in these ecosystems. The results of this study provide a scientific basis for relevant departments to formulate future policies addressing the impact of extreme climate on vegetation in different ecosystems on the QXP.
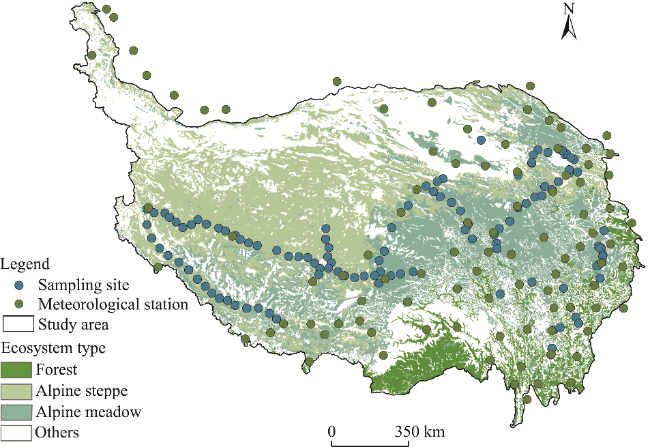
Fig. 1 Overview of the Qinghai-Xizang Plateau (QXP) and distributions of meteorological stations and sampling sites. Ecosystem type data are based on the vegetation type data sourced from the Resource and Environmental Science Data Platform (https://www.resdc.cn/data.aspx?DATAID=122). |
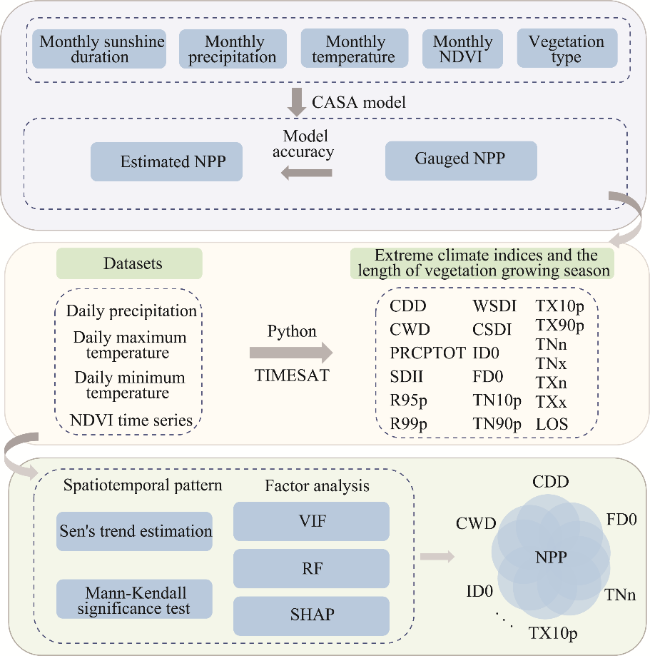
Fig. 2 Flowchart of this study. NDVI, normalized difference vegetation index; CASA, Carnegie-Ames-Stanford Approach; NPP, net primary productivity; CDD, consecutive dry days; CWD, consecutive wet days; PRCPTOT, total wet-day precipitation; SDII, simple daily intensity index; R95p, very-wet-day precipitation; R99p, extremely-wet-day precipitation; WSDI, warm spell duration index; CSDI, cold spell duration index; ID0, ice days; FD0, frost days; TN10p, cool nights; TN90p, warm nights; TX10p, cool days; TX90p, warm days; TNn, coldest night; TNx, warmest night; TXn, coldest day; TXx, hottest day; LOS, length of vegetation growing season; VIF, Variance Inflation Factor; RF, random forest; SHAP, SHapley Additive exPlanations. |
Table 1 Definitions of extreme precipitation indices |
| Index | Unit | Description |
|---|---|---|
| Consecutive dry days (CDD) | d | Maximum number of consecutive days with daily precipitation <1 mm |
| Consecutive wet days (CWD) | d | Maximum number of consecutive days with daily precipitation ≥1 mm |
| Total wet-day precipitation (PRCPTOT) | mm | Annual total precipitation on wet days |
| Simple daily intensity index (SDII) | mm/d | Annual total precipitation divided by the number of wet days in the year |
| Very-wet-day precipitation (R95p) | mm | Annual total precipitation of days with daily precipitation >95th percentile |
| Extremely-wet-day precipitation (R99p) | mm | Annual total precipitation of days with daily precipitation >99th percentile |
Note: 95th percentile and 99th percentile represent 95th percentile and 99th percentile of precipitation on wet days for the period 1982-2020, respectively. Precise definitions are given at https://etccdi.pacificclimate.org/list_27_indices.shtml. |
Table 2 Definitions of extreme temperature indices |
| Index | Unit | Description |
|---|---|---|
| Warm spell duration index (WSDI) | d | Number of days with at least six consecutive days when TX>90th percentile |
| Cold spell duration index (CSDI) | d | Number of days with at least six consecutive days when TN<10th percentile |
| Ice days (ID0) | d | Number of days when TX<0°C |
| Frost days (FD0) | d | Number of days when TN<0°C |
| Cool nights (TN10p) | % | Percentage of days when TN<10th percentile |
| Warm nights (TN90p) | % | Percentage of days when TN>90th percentile |
| Cool days (TX10p) | % | Percentage of days when TX<10th percentile |
| Warm days (TX90p) | % | Percentage of days when TX>90th percentile |
| Coldest night (TNn) | °C | Minimum of TN |
| Warmest night (TNx) | °C | Maximum of TN |
| Coldest day (TXn) | °C | Minimum of TX |
| Hottest day (TXx) | °C | Maximum of TX |
Note: TX and TN denote the daily maximum temperature and daily minimum temperature, respectively. The 10th percentile denotes the calendar day 10th percentile centred on a 5-d window for the base period 1982-2020. The 90th percentile denotes the calendar day 90th percentile centred on a 5-d window for the base period 1982-2020. Precise definitions are given at https://etccdi.pacificclimate.org/ list_27_indices.shtml. |
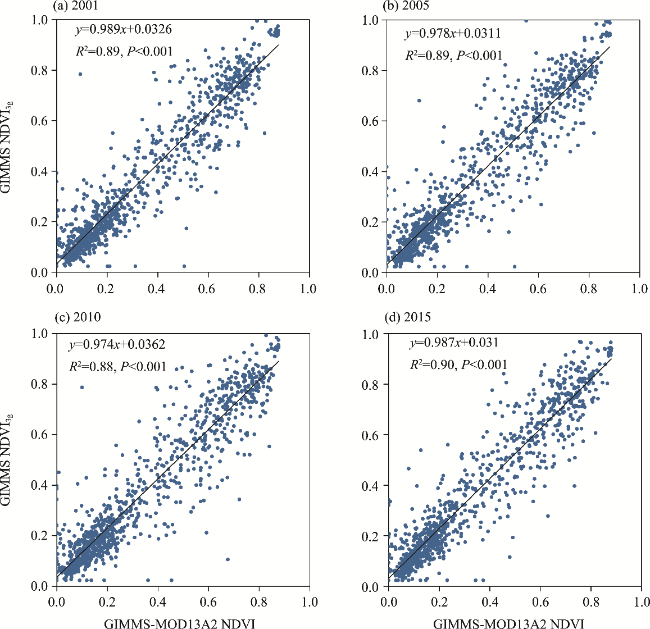
Fig. 3 Linear relationship between the GIMMS-MOD13A2 NDVI and GIMMS NDVI3g in 2001 (a), 2005 (b), 2010 (c), and 2015 (d). GIMMS, Global Inventory Modeling and Mapping Studies. GIMMS-MOD13A2 NDVI is the simulated NDVI based on MOD13A2 NDVI and GIMMS NDVI3g datasets. |
Fig. 4 Linear relationship between the gauged and estimated NPP data. r, BIAS, ME, and RMSE denote the correlation coefficient, relative bias, mean error, and root mean square error, respectively. |
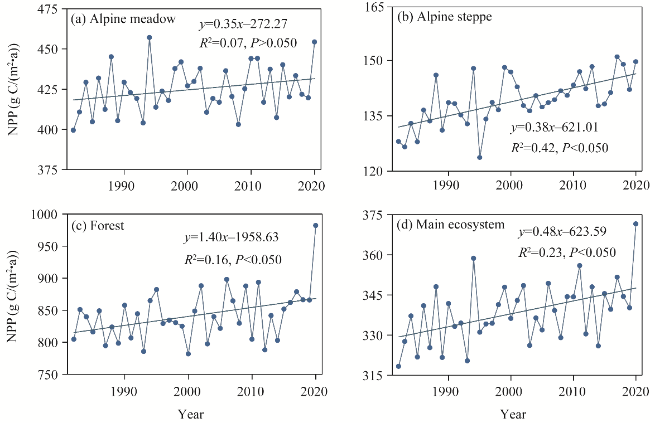
Fig. 5 Temporal variations in NPP in different ecosystems on the QXP from 1982 to 2020. (a), alpine meadow; (b), alpine steppe; (c), forest; (d), main ecosystem. |
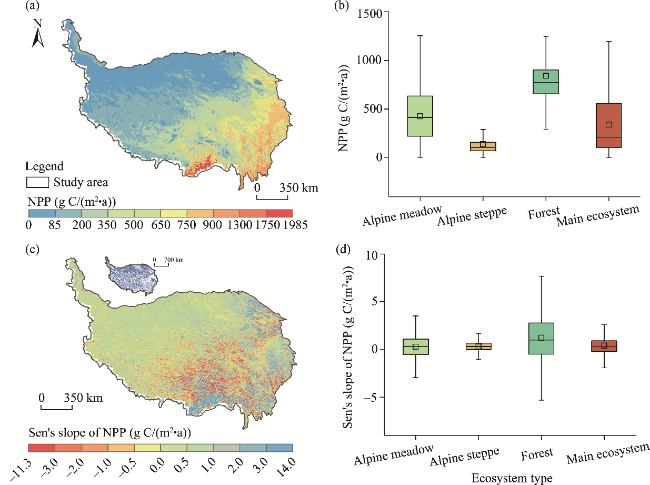
Fig. 6 Spatial patterns and zonal statistics of multi-year average NPP (a and b) and Sen's slope of NPP (c and d) on the QXP during 1982-2020. The inset in the upper left corner of Figure 6c shows that the colored pixels experienced significant changes (P<0.050 level). In the box plots, the box boundaries denote the 25th and 75th percentiles, the line in the box denotes the median, the square in the box denotes the average value, and whiskers below and above the box denote the Q1-1.5(Q3-Q1) and Q3+1.5(Q3-Q1). |
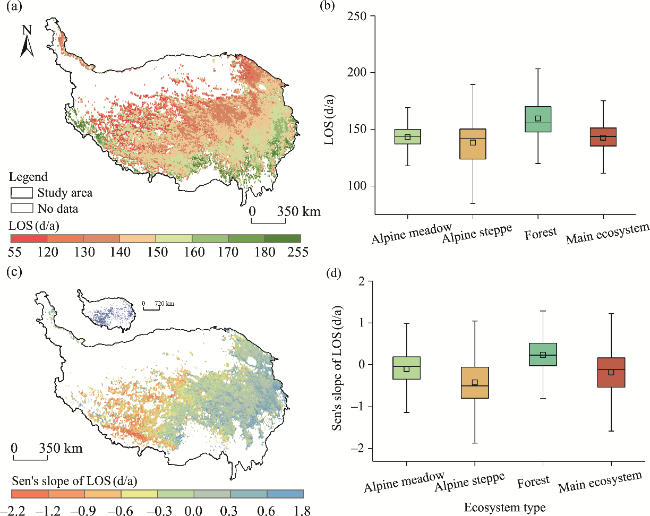
Fig. 7 Spatial patterns and zonal statistics of the multi-year average LOS (a and b) and Sen's slope of LOS (c and d) on the QXP during 1982-2020. The inset in the upper left corner of Figure 7c shows that the colored pixels experienced significant changes (P<0.050 level). In the box plots, the box boundaries denote the 25th and 75th percentiles, the line in the box denotes the median, the square in the box denotes the average value, and whiskers below and above the box denote the Q1-1.5(Q3-Q1) and Q3+1.5(Q3-Q1). |
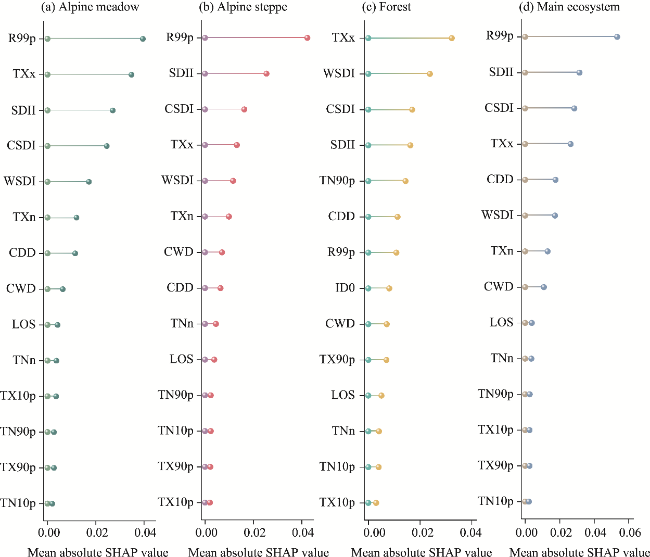
Fig. 8 Relative importances of extreme climate indices and the length of vegetation growing season based on the mean absolute SHAP values |
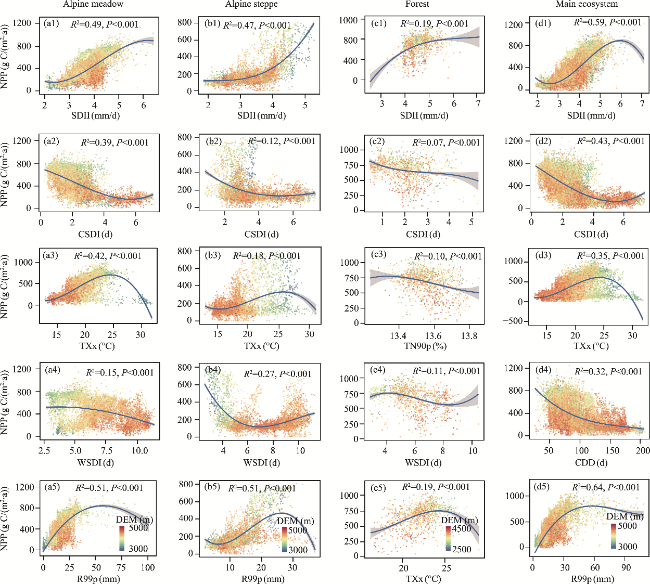
Fig. 9 Relationships between the NPP and extreme climate indices in the alpine meadow (a1-a5), alpine steppe (b1-b5), forest (c1-c5), and main ecosystem (d1-d5). The selection of extreme climate indices for different ecosystems is based on the mean absolute SHAP values. The DEM legend applies to all scatter plots within its respective column. The shaded area denotes the 95% confidence interval of the curve fitting. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}