
Prospects and limitations of soil amendment and irrigation techniques for the water-saving public urban greenery and ephemeral weed management in the sandy soils of the United Arab Emirates
The second and the first authors contributed equally to this work.
Received date: 2024-04-08
Revised date: 2024-07-31
Accepted date: 2024-08-23
Online published: 2025-08-13
Ayesha ALAM , Elke GABRIEL-NEUMANN . [J]. Journal of Arid Land, 2024 , 16(9) : 1288 -1302 . DOI: 10.1007/s40333-024-0028-9
Public urban greenery greatly contributes to the residential and tourist value of cities in the Gulf Region, but due to the hyper-arid climatic conditions, the cost of irrigation and plant maintenance is very high. Existing strategies to reduce the monetary and ecological costs involve the cultivation of native xerophytic plantations, and/or the use of soil improvers to increase water- and nutrient-holding capacity of the sandy soils. Various soil improvers based on mineral, organic, or synthetic materials have entered the United Arab Emirates (UAE) market in recent years, but there is considerable uncertainty about how they should best be used in combination with ornamental plant stands involving xerophytic native plants. The present study investigated the effect of soil amendment and deep pipe irrigation on perennial ornamental plant stands involving native plants (Tephrosia appolinea (Gel.) Link in combination with Aerva javanica (Burm. f.) Juss. ex Schult.) and native-exotic plants (T. appolinea in combination with Ruelia simplex C. Wright) either or not topsoil and subsoil amendment with bentonite and hydrophobic sand under the irrigation water supply of less than 50% of reference evapotranspiration (ET0). After one year of cultivation, T. appolinea and A. javanica (native vs. native) produced high biomass and exhibited high water use efficiency (WUE) as compared with T. appolinea and R. simplex (native vs. exotic) combination given that no significant differences were found under the soil amendment treatments. All stands thrived under irrigation water supply far below what is usually supplied to exotic ornamental stands in public parks of the Al Ain City, the UAE. However, subsoil amendment in combination with deep pipe irrigation reduced the occurrence of weeds and increased the overall plant rooting depth. Our results suggest that subsoil amendment and irrigation up to 60-80 cm depth can potentially control ephemeral weed infestation, which is a great challenge in various plant production systems of the Gulf Region. The results of the present study suggest that the impact of soil amendment on the WUE of exotic plants is marginal and might not be economically justified. Replacing exotic with native ornamental plant species seems to have a far greater water-saving potential than the amendment of the soil, while weeds can be suppressed in the absence of topsoil moisture.
Table 1 Experimental design indicating different plant combination, soil amendment, and irrigation treatments |
| Treatment 1 | Treatment 2 | Treatment 3 | Treatment 4 | ||||
|---|---|---|---|---|---|---|---|
| Control | Topsoil amendment | Subsoil amendment with hydrophobic sand | Subsoil amendment with deep pipe irrigation | ||||
| No amendment | Potash bentonite with 10 kg/m2 rate at 25 cm depth | A 1-2-cm thick layer of hydrophobic sand at a depth of 60 cm | A 1-2-cm thick layer of hydrophobic sand at a depth of 60 cm of soil, 75 cm long deep pipes, and potash bentonite with 10 kg/m2 rate at 40-50 cm depth | ||||
| native vs. native | native vs. exotic | native vs. native | native vs. exotic | native vs. native | native vs. exotic | native vs. native | native vs. exotic |
Note: Total: 56 plant plots. |
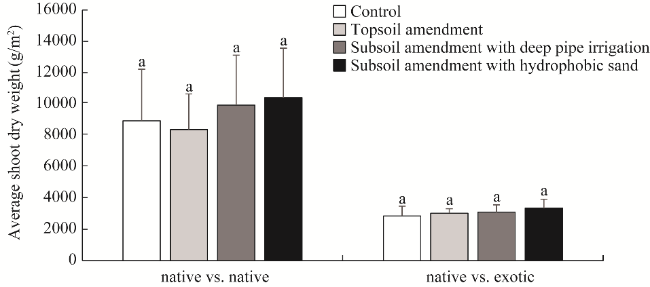
Fig. 1 Average shoot dry weights of plants in native vs. native and native vs. exotic plots under different treatments. Different lowercase letters within the same plot indicate significant differences at P<0.05 level among different treatments. |
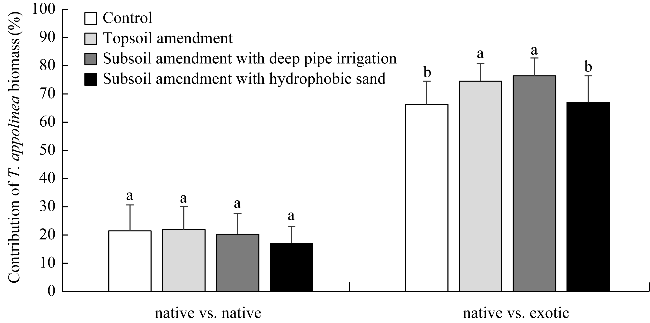
Fig. 2 Contribution of Tephrosia appolinea (Gel.) Link biomass in total biomass from native vs. native and native vs. exotic plots under different treatments. Different lowercase letters within the same plot indicate significant differences at P<0.05 level among different treatments. |
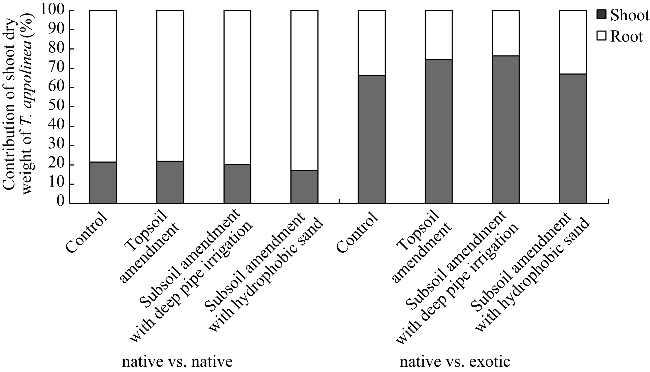
Fig. 3 Contribution of shoot dry weight of T. appolinea in total weights from native vs. native and native vs. exotic plots under different treatments |
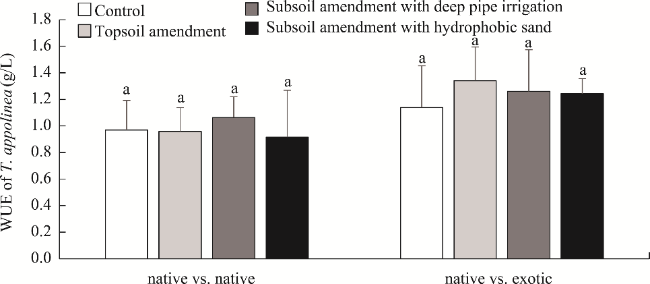
Fig. 4 Water use efficiency (WUE) of T. appolinea from native vs. native and native vs. exotic plots under different treatments. Different lowercase letters within the same plot indicate significant difference at P<0.05 level among different treatments. |
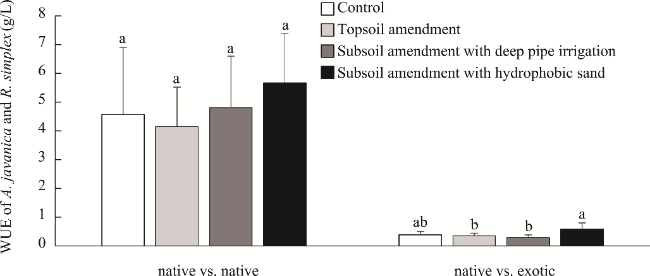
Fig. 5 WUE of Aerva javanica (Burm. f.) Juss. ex Schult. and Ruelia simplex C. Wright from native vs. native and native vs. exotic plots under different treatments. Different lowercase letters within the same plot indicates significant differences at P<0.05 level among different treatments. |
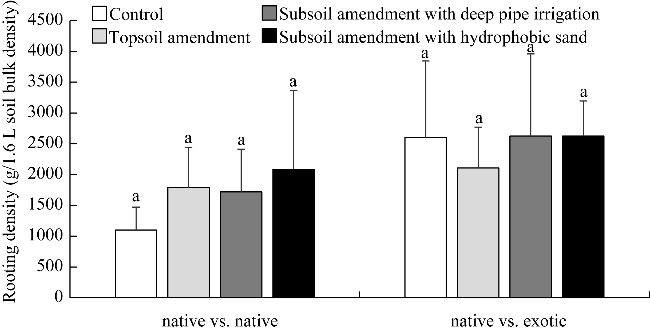
Fig. 6 Rooting density of plants from native vs. native and native vs. exotic plots under different treatments. Different lowercase letters within the same plot indicate significant differences at P<0.05 level among different treatments. |
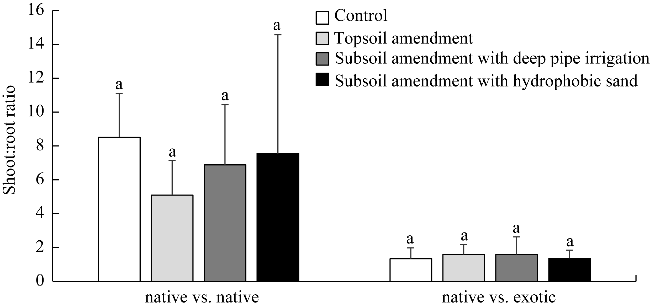
Fig. 7 Shoot:root ratio of plants from native vs native and native vs exotic plots under different treatments. Different lowercase letters within the same plot indicate significant differences at P<0.05 level among different treatments. |
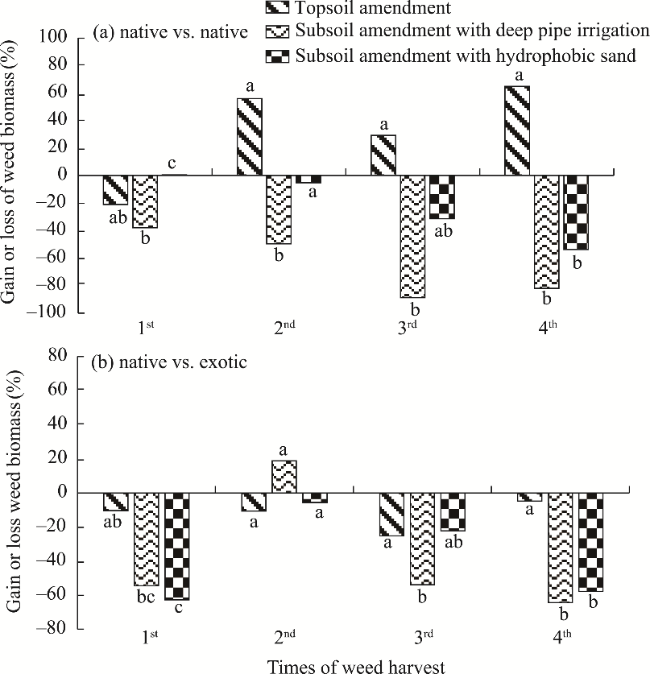
Fig. 8 Gain (bars above the axis line) or loss (bar below the axis line) in weed biomass during four times of harvests under different treatments, as compared with the control in native vs. native plot (a) and native vs. exotic plot (b). Different lowercase letters indicate significant differences at P<0.05 level among different times of weed harvest and treatments. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
FAO (Food and Agriculture Organization of the United Nations). 2020. Crop production manual guide to fruit and vegetable production in the Federated States of Micronesia. [2024-01-28]. https://openknowledge.fao.org/server/api/core/bitstreams/aac462ae-90d2-422c-b9e6-3e5336b18b52/content.
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
Masdar. 2019. Sustainable Agriculture for Arid Climates. [2023-12-09]. http://www.scribd.com/document/564847626/Sustainable-agriculture-for-aird-climate.
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}