
Thriving green havens in baking deserts: Plant diversity and species composition of urban plantations in the Sahara Desert
Received date: 2024-01-17
Revised date: 2024-06-27
Accepted date: 2024-06-28
Online published: 2025-08-13
Mohammed SOUDDI , Haroun CHENCHOUNI , M'hammed BOUALLALA . [J]. Journal of Arid Land, 2024 , 16(9) : 1270 -1287 . DOI: 10.1007/s40333-024-0083-2
Hot arid zones represent vital reservoirs of unique species and ecosystems, holding significant importance for biodiversity. This study aimed to explore the plant diversity associated with tree plantations in urban ecosystems under hyper-arid climatic conditions in the Sahara Desert of Algeria. In May 2022, 30 quadrats measuring 1 m2 each were established at the base of Phoenix dactylifera, Leucaena leucocephala, and Tamarix aphylla, corresponding to the dominant tree species in each of three plantations. In each quadrat, the plant quantitative inventory was conducted to measure plant diversity and similarity among the studied plantations. Based on this, we assessed the plant functional traits and rarity/abundance status of the flora. The findings revealed a diverse flora associated with the studied plantations, comprising 29 plant species grouped into 27 genera and 12 families. Notably, Poaceae (accounting for 30.8% of the flora), Asteraceae (25.0%), and Zygophyllaceae (21.6%) were well-represented. With an overall density of approximately 555 individuals/m2, Zygophyllum album (120 individuals/m2) and Polypogon monspeliensis (87 individuals/m2) emerged as the most abundant species. Functional trait analysis underscored the pivotal role of therophytes (constituting over 50.0% of the flora) and anemochorous species (33.0%-62.5%). Phytogeographic analysis emphasized the prevalence of the Saharo-Arabic element (constituting over 31.0% of the flora) and the Mediterranean Saharo-Arabic element (9.5%-21.5%). The Cosmopolitan element thrived under disturbance factors, recording percentages from 13.0% to 20.0% of the plant community. The rarity/abundance status of the flora emphasized the significance of rare, common, and very common species in the studied plantations. These findings could provide fundamental data for the effective control and management of biodiversity in hot hyper-arid urban ecosystems.
Table 1 Plant density, plant functional traits, and rarity/abundance status of plant species associated to the three urban plantations in the Sahara Desert of Algeria |
| Family | Species | n | LF | MT | DT | CT | AA | Occ (%) | Class Occ | Plantations |
|---|---|---|---|---|---|---|---|---|---|---|
| Amaranthaceae | Bassia muricata | 56 | Ther | Annual | Zoo | M-SA | C | 56.7 | III | Lle+Pda+Tap |
| Chenopodium murale | 17 | Ther | Annual | Baro | Cosm | R | 16.7 | I | Lle+Pda+Tap | |
| Salsola foetida | 2 | Cham | Perennial | Anemo | SA | R | 3.3 | I | Pda | |
| Apocynaceae | Calotropis procera | 18 | Phan | Perennial | Anemo | Tr-SA | R | 36.7 | II | Lle+Pda+Tap |
| Cynanchum acutum | 1 | Hemi | Perennial | Anemo | M-As | RR | 3.3 | I | Pda | |
| Pergularia tomentosa | 5 | Cham | Perennial | Anemo | SA | CC | 13.3 | I | Lle+Pda | |
| Asteraceae | Aster squamatus | 2 | Ther | Annual | Anemo | S-Am | C | 6.7 | I | Lle+Pda |
| Lactuca serriola | 14 | Ther | Annual | Anemo | PTm | AC | 20.0 | I | Pda+Tap | |
| Launaea glomerata | 16 | Ther | Annual | Anemo | M-SA | RR | 30.0 | II | Lle+Pda+Tap | |
| Launaea nudicaulis | 51 | Ther | Annual | Anemo | SA | CC | 53.3 | III | Lle+Pda+Tap | |
| Launaea resedifolia | 2 | Ther | Annual | Anemo | M-SA | CC | 3.3 | I | Pda | |
| Pulicaria arabica | 15 | Hemi | Perennial | Anemo | SA | AC | 10.0 | I | Pda+Tap | |
| Senecio massaicus | 7 | Ther | Annual | Anemo | SA | R | 3.3 | I | Pda | |
| Sonchus oleraceus | 32 | Ther | Annual | Anemo | Cosm | CCC | 36.7 | II | Lle+Pda+Tap | |
| Convovulaceae | Merremia dissecta | 2 | Ther | Annual | Anthropo | Tr-Am | - | 6.7 | I | Lle+Pda |
| Cucurbitaceae | Colocynthis vulgaris | 1 | Ther | Annual | Anemo | M-SA | CC | 3.3 | I | Lle |
| Euphorbiaceae | Euphorbia granulate | 1 | Ther | Annual | Baro | SA | C | 3.3 | I | Pda |
| Heliotropiaceae | Heliotropium bacciferum | 7 | Cham | Perennial | Baro | SA | CC | 16.7 | I | Lle+Pda+Tap |
| Orobanchaceae | Cistanche phelypaea | 6 | Ther | Annual | Anemo | M-SA | C | 6.7 | I | Lle+Tap |
| Poaceae | Cutandia dichotoma | 9 | Ther | Annual | Anemo | M | C | 16.7 | I | Lle+Pda |
| Cynodon dactylon | 5 | Hemi | Perennial | Baro | Cosm | CCC | 3.3 | I | Pda | |
| Imperata cylindrica | 20 | Hemi | Perennial | Anemo | Tr-M-SA | AC | 3.3 | I | Pda | |
| Lolium multiflorum | 29 | Ther | Annual | Baro | M | CC | 33.3 | II | Lle+Pda+Tap | |
| Phalaris minor | 1 | Ther | Annual | Zoo | PSTr | AR | 3.3 | I | Lle | |
| Phragmites australis | 20 | Hemi | Perennial | Anemo | Cosm | C | 16.7 | I | Lle+Pda | |
| Polypogon monspeliensis | 87 | Ther | Annual | Zoo | PSTr | CC | 36.7 | II | Lle+Pda+Tap | |
| Primulaceae | Anagallis arvensis | 1 | Ther | Annual | Anemo | Cosm | R | 3.3 | I | Tap |
| Resedaceae | Randonia africana | 8 | Cham | Perennial | Baro | SA | R | 16.7 | I | Lle+Tap |
| Zygophyllaceae | Zygophyllum album | 120 | Cham | Perennial | Baro | SA | C | 66.7 | IV | Lle+Pda+Tap |
Note: n, the number of individuals of a species; LF, life form; Ther, therophyte; Cham, chamaephyte; Phan, phanerophyte; Hemi, hemicryptophytes; MT, morphological type; DT, dispersal type; Zoo, zoochore; Baro, barochore; Anemo, anemochore; Anthropo, anthropochore; CT, chorological type; M-SA, Mediterranean Saharo-Arabian; Cosm, Cosmopolitan; SA, Saharo-Arabian; Tr-SA, Tropico-Saharo-Arabian; M-As, Mediterrano-Asian; S-Am, South American; PTm, Paleo-temperate; Tr-Am, Tropico-American; M, Mediterranean; Tr-M-SA, Tropico-Mediterranean Saharo-Arabian; PSTr, Paleo-subtropical; AA, abundance appraisal; C, common; R, rare; RR, very rare; CC, very common; AC, fairly common; CCC, particularly widespread; AR, quite rare; Occ, frequency of occurrence; Class Occ, the class for the frequency of occurrence. Class I is very rare species with Occ<20.0%; Class II is rare species with Occ varying from 20.0% to 40.0%; Class III is frequent species with Occ varying from 40.0% to 60.0%; Class IV is abundant species with Occ varying from 60.0%-80.0%. Pda, Phoenix dactylifera; Lle, Leucaena leucocephala; Tap, Tamarix aphylla. "-" means no category. |
Table 2 Genus richness and species richness for different plant families identified in the three urban plantations in the Sahara Desert of Algeria |
| Family | Pda plantation | Lle plantation | Tap plantation | Overall | ||||
|---|---|---|---|---|---|---|---|---|
| Genus richness | Species richness | Genus richness | Species richness | Genus richness | Species richness | Genus richness | Species richness | |
| Amaranthaceae | 3 (13.6%) | 3 (12.5%) | 2 (11.1%) | 2 (10.5%) | 2 (14.3%) | 2 (13.3%) | 3 (11.1%) | 3 (10.3%) |
| Apocynaceae | 3 (13.6%) | 3 (12.5%) | 2 (11.1%) | 2 (10.5%) | 1 (7.1%) | 1 (6.7%) | 3 (11.1%) | 3 (10.3%) |
| Asteraceae | 6 (27.3%) | 8 (33.3%) | 3 (16.7%) | 4 (21.1%) | 4 (28.6%) | 5 (33.3%) | 6 (22.2%) | 8 (27.6%) |
| Convovulaceae | 1 (4.5%) | 1 (4.2%) | 1 (5.6%) | 1 (5.3%) | - | - | 1 (3.7%) | 1 (3.4%) |
| Cucurbitaceae | - | - | 1 (5.6%) | 1 (5.3%) | - | - | 1 (3.7%) | 1 (3.4%) |
| Euphorbiaceae | 1 (4.5%) | 1 (4.2%) | - | - | - | - | 1 (3.7%) | 1 (3.4%) |
| Heliotropiaceae | 1 (4.5%) | 1 (4.2%) | 1 (5.6%) | 1 (5.3%) | 1 (7.1%) | 1 (6.7%) | 1 (3.7%) | 1 (3.4%) |
| Orobanchaceae | - | - | 1 (5.6%) | 1 (5.3%) | 1 (7.1%) | 1 (6.7%) | 1 (3.7%) | 1 (3.4%) |
| Poaceae | 6 (27.3%) | 6 (25.0%) | 5 (27.8%) | 5 (26.3%) | 2 (14.3%) | 2 (13.3%) | 7 (25.9%) | 7 (24.1%) |
| Primulaceae | - | - | - | - | 1 (7.1%) | 1 (6.7%) | 1 (3.7%) | 1 (3.4%) |
| Resedaceae | - | - | 1 (5.6%) | 1 (5.3%) | 1 (7.1%) | 1 (6.7%) | 1 (3.7%) | 1 (3.4%) |
| Zygophyllaceae | 1 (4.5%) | 1 (4.2%) | 1 (5.6%) | 1 (5.3%) | 1 (7.1%) | 1 (6.7%) | 1 (3.7%) | 1 (3.4%) |
| Total | 22 (100.0%) | 24 (100.0%) | 18 (100.0%) | 19 (100.0%) | 14 (100.0%) | 15 (100.0%) | 27 (100.0%) | 29 (100.0%) |
Note: "-" means no value. The values in parentheses are the corresponding percentages. |
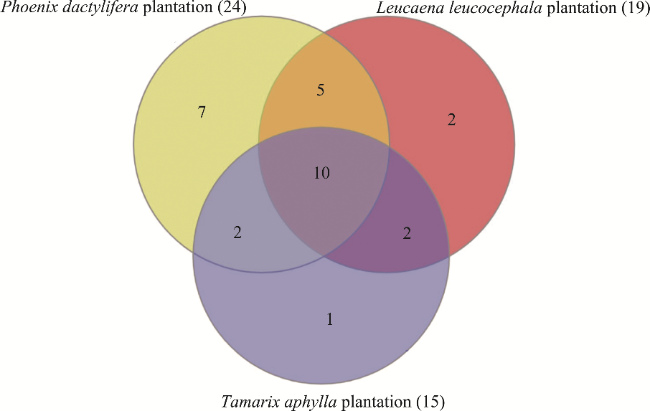
Fig. 1 Venn diagram illustrating the partitioning of plant species richness in the three urban plantations in the Sahara Desert of Algeria |
Table 3 Qualitative and quantitative similarity estimates of plant community composition between each two urban plantations in the Sahara Desert of Algeria |
| Estimate index | Lle-Pda plantations | Lle-Tap plantations | Pda-Tap plantations |
|---|---|---|---|
| Number of shared species observed in the two plantations | 15 | 12 | 12 |
| Estimated number of Chao-shared species | 18.5 | 12.3 | 12.1 |
| Classic Jaccard index (%) | 53.6 | 54.5 | 44.4 |
| Classic Sørensen index (%) | 69.8 | 70.6 | 61.5 |
| Raw Chao-Jaccard index (%) | 70.5 | 82.2 | 65.6 |
| Estimated Chao-Jaccard index (%) | 73.0 | 83.2 | 66.0 |
| Raw Chao-Sørensen index (%) | 82.7 | 90.2 | 79.2 |
| Estimated Chao-Sørensen index (%) | 84.4 | 90.9 | 79.5 |
| Morisita-Horn index (%) | 73.1 | 78.1 | 79.0 |
| Bray-Curtis index (%) | 59.0 | 63.8 | 61.5 |
Table 4 Plant diversity indices of plant communities associated with the three urban plantations in the Sahara Desert of Algeria |
| Diversity index | Pda plantation | Lle plantation | Tap plantation |
|---|---|---|---|
| Species richness | 24 | 19 | 15 |
| Shannon-Wiener index | 2.73 | 2.26 | 2.35 |
| Equitability index | 0.86 | 0.77 | 0.87 |
| Chao-1 index | 29 | 24 | 15 |
| Simpson's diversity index | 0.91 | 0.84 | 0.88 |
| Brillouin index | 2.52 | 2.10 | 2.21 |
| Margalef index | 4.41 | 3.46 | 2.67 |
| Berger-Parker index | 0.16 | 0.34 | 0.21 |
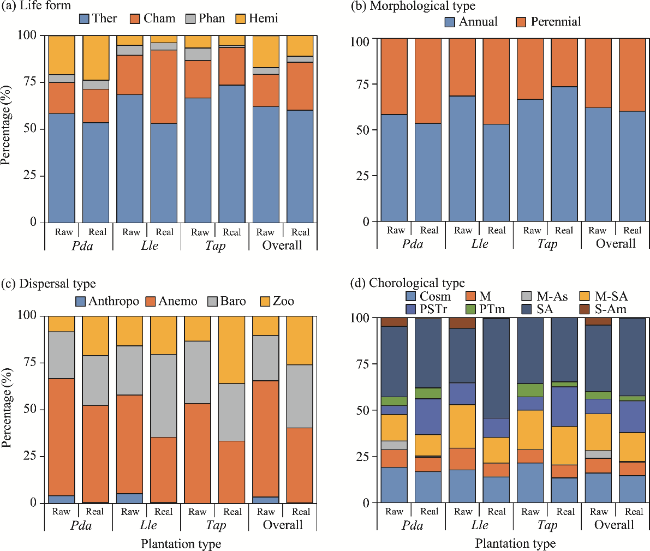
Fig. 2 Distribution of different life forms (a), morphological types (b), dispersal types (c), and chorological types (d) identified in the three urban plantations in the Sahara Desert of Algeria in both raw and real spectra. Pda, Phoenix dactylifera; Lle, Leucaena leucocephala; Tap, Tamarix aphylla. Ther, therophyte; Cham, chamaephyte; Phan, phanerophyte; Hemi, hemicryptophytes; Anthropo, anthropochore; Anemo, anemochore; Baro, barochore; Zoo, zoochore; Cosm, Cosmopolitan; Med, Mediterranean; M-As, Mediterrano-Asian; M-SA, Mediterranean Saharo-Arabian; PSTr, Paleo-subtropical; PTm, Paleo-temperate; SA, Saharo-Arabian; S-Am, South American. |
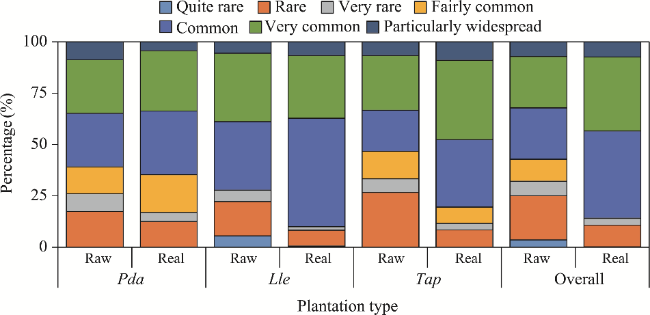
Fig. 3 Rarity/abundance status of the flora in the three urban plantations in the Sahara Desert of Algeria in both raw and real spectra |
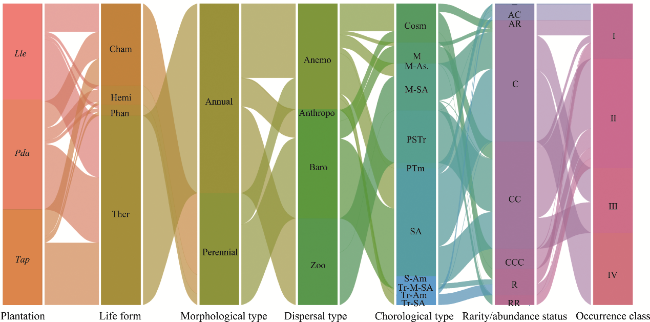
Fig. 4 Alluvial diagram displaying the distribution of life forms, morphological types, dispersal types, chorological types, rarity/abundance status, and occurrence classes of the three urban plantations in the Sahara Desert of Algeria. Tr-M-SA, Tropico-Mediterranean Saharo-Arabian; Tr-Am, Tropico-American; Tr-SA, Tropico-Saharo-Arabian; AR, quite rare; R, rare; RR, very rare; AC, fairly common; C, common; CC, very common; CCC, particularly widespread. Class I is very rare species with frequency of occurrence<20.0%; Class II is rare species with frequency of occurrence varying from 20.0% to 40.0%; Class III is frequent species with frequency of occurrence varying from 40.0% to 60.0%; Class IV is abundant species with frequency of occurrence varying from 60.0%-80.0%. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
FAO (Food and Agriculture Organization of the United Nations). 2019. Trees, forests and land use in drylands: the first global assessment-Full report. FAO Forestry Paper 184. FAO, Rome, Italy.
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
MEA (Millennium Ecosystem Assessment). 2005. Ecosystem Wealth and Human Well-being. Washington DC: Island Press, 1-135.
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
Roselt/OSS (Observatory Network for Long-Term Ecological Monitoring). 2008. Methodological guide for the study and monitoring of flora and vegetation. Tunis: Sahara and Sahel Observatory, 171. (in French)
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}