
Mechanical properties and enhanced soil shear strength of herbaceous plant roots in the alpine meadow layer of the permafrost region on the Qinghai-Xizang Plateau, China
The second and third authors contributed equally to this work.
Received date: 2024-11-21
Revised date: 2025-03-10
Accepted date: 2025-03-17
Online published: 2025-08-13
HE Dequan , LU Haijing , HU Xiasong , WANG Cheng , LIU Changyi , ZHAO Yingxiao , LI Shuaifei , DENG Taiguo . [J]. Journal of Arid Land, 2025 , 17(4) : 515 -537 . DOI: 10.1007/s40333-025-0051-5
The Qinghai-Xizang Plateau of China faces challenges like thaw slumping, threatening slope stability and infrastructure. Understanding the mechanical properties of the roots of the dominant herbaceous plant species in the alpine meadow layer of the permafrost regions on the Qinghai-Xizang Plateau is essential for evaluating their role in enhancing soil shear strength and mitigating slope deformation in these fragile environments. In this study, the roots of four dominant herbaceous plant species—Kobresia pygmaea, Kobresia humilis, Carex moorcroftii, and Leontopodium pusillum—that are widely distributed in the permafrost regions of the Qinghai-Xizang Plateau were explored to determine their mechanical properties and effects in enhancing soil shear strength. Through indoor single root tensile and root group tensile tests, we determined the root diameter, tensile force, tensile strength, tensile ratio, and strength frequency distributions. We also evaluated their contributions to inhibiting slope deformation and failure during the formation and development of thermal thaw slumps in the alpine meadow. The results showed that the distribution of the root diameter of the dominant plant species is mostly normal, while the tensile strength tends to be logarithmically normally distributed. The relationship between the root diameter and root tensile strength conforms to a power function. The theoretical tensile strength of the root group was calculated using the Wu-Waldron Model (WWM) and the Fiber Bundle Model (FBM) under the assumption that the cumulative single tensile strength of the root bundle is identical to the tensile strength of the root group in the WWM. The FBM considers three fracture modes: FBM-D (the tensile force on each single root is proportional to its diameter relative to the total sum of all the root diameters), FBM-S (the cross-sectional stress in the root bundle is uniform), and FBM-N (each tensile strength test of individual roots experiences an equal load). It was found that the model-calculated tensile strength of the root group was 162.60% higher than the test value. The model-derived tensile force of the root group from the FBM-D, FBM-S, and FBM-N was 73.10%, 28.91%, and 13.47% higher than the test values, respectively. The additional cohesion of the soil provided by the roots was calculated to be 25.90-45.06 kPa using the modified WWM, 67.05-38.15 kPa using the FBM-S, and 57.24-32.74 kPa using the FBM-N. These results not only provide a theoretical basis for further quantitative evaluation of the mechanical effects of the root systems of herbaceous plant species in reinforcing the surface soil but also have practical significance for the effective prevention and control of thermal thaw slumping disasters in the permafrost regions containing native alpine meadows on the Qinghai-Xizang Plateau using flexible plant protection measures.
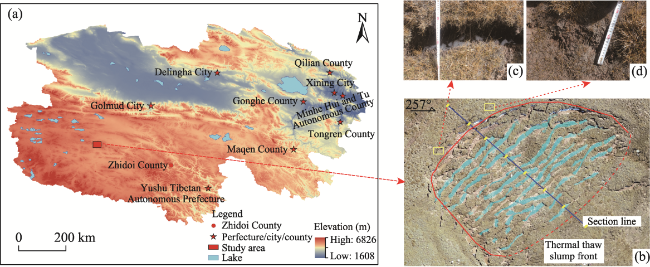
Fig. 1 Location of the study area (a) and development range and tensile crack distribution characteristics of the selected thermal thaw slump slope, including (b) thermal thaw slumping distribution area, (c) formation of tensile cracks (4 cm depth), and (d) continuous tensile cracks (10 cm depth) |
Fig. 2 Photos showing field sampling, root sample preparation, and testing procedure. (a), field sampling; (b), root preparation; (c), root samples; (d), universal testing machine; (e), tensile test process; (f), group root before tensile test; (g), group root after tensile test. |
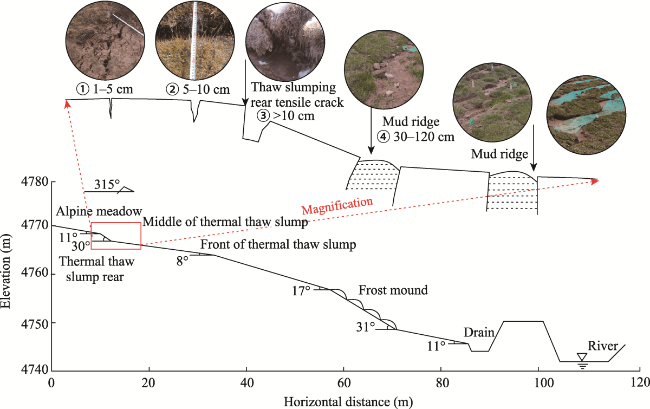
Fig. 3 Development of tensile cracks at the trailing edge of thermal thaw slump slope in the study area, along with the tensile crack characteristics at different development stages. This figure is modified from the work of Wang et al. (2024b) and supplemented with additional relevant results from this study. |
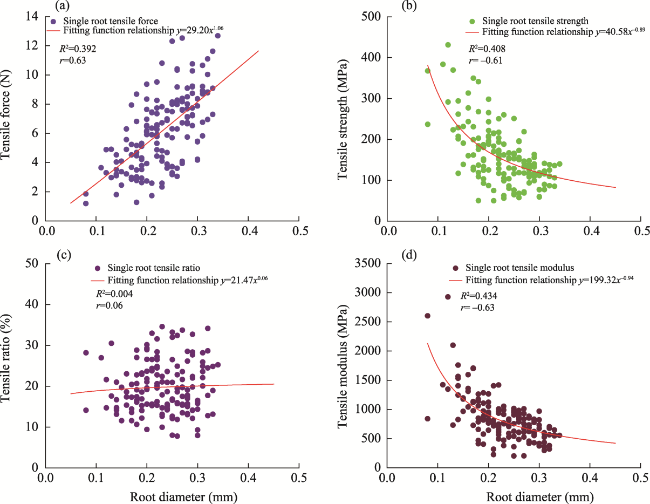
Fig. 4 Tensile test results of individual roots in the alpine meadow layer of the study area and their fitting relationships with root diameter. (a), root diameter versus single root tensile force; (b), root diameter versus single root tensile strength; (c), root diameter versus single root tensile ratio; (d), root diameter versus single root tensile modulus. R2 is the goodness of fit, and r is the Pearson correlation coefficient. |
Table 1 Test results of root diameter and single root tensile properties for the dominant plant species in the alpine meadow layer of the study area |
| Statistic | Root diameter (mm) | Singe root tensile force (N) | Singe root tensile strength (MPa) | Singe root tensile ratio (%) | Singe root tensile modulus (MPa) |
|---|---|---|---|---|---|
| Minimum | 0.08 | 1.19 | 49.90 | 7.79 | 201.88 |
| Maximum | 0.34 | 12.68 | 431.20 | 34.59 | 2928.35 |
| Median | 0.23 | 6.30 | 140.50 | 19.11 | 781.43 |
| Average | 0.23 | 6.19 | 158.19 | 19.74 | 841.85 |
| Standard deviation | 0.06 | 2.54 | 71.63 | 6.16 | 394.75 |
| Coefficient of variation | 24.46 | 40.96 | 45.28 | 31.18 | 46.89 |
| Skew coefficient | -0.28 | 0.25 | 1.19 | 0.45 | 1.94 |
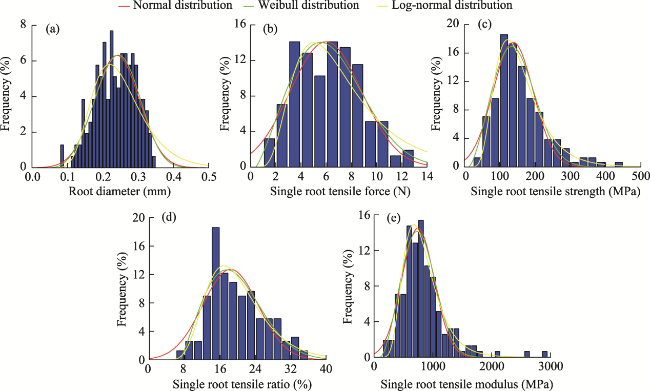
Fig. 5 Probability density histograms and fitting curves for root diameter and single root tensile properties of dominant plant species in the alpine meadow layer of the study area. (a), root diameter; (b), single root tensile force; (c), single root tensile strength; (d), single root tensile ratio; (e), single root tensile modulus. |
Table 2 Fitting results of different distribution functions for root diameter and single root tensile properties of dominant plant species in the alpine meadow layer of the study area |
| Fitting function | Fitting function parameter | Root diameter | Singe root tensile force | Singe root tensile strength | Singe root tensile ratio | Singe root tensile modulus |
|---|---|---|---|---|---|---|
| Normal distribution | μ | 0.24 | 5.97 | 138.28 | 18.36 | 746.83 |
| σ | 0.07 | 2.83 | 56.96 | 6.28 | 275.35 | |
| R2 | 0.813 | 0.858 | 0.932 | 0.663 | 0.940 | |
| Weibull distribution | α | 21.83 | 6.67 | 5.97 | 7.09 | 7.55 |
| β | 3.60 | 2.26 | 2.50 | 2.14 | 2.68 | |
| u | 4.49 | 0.40 | 0.50 | 3.49 | 1.00 | |
| R2 | 0.809 | 0.875 | 0.943 | 0.705 | 0.93 | |
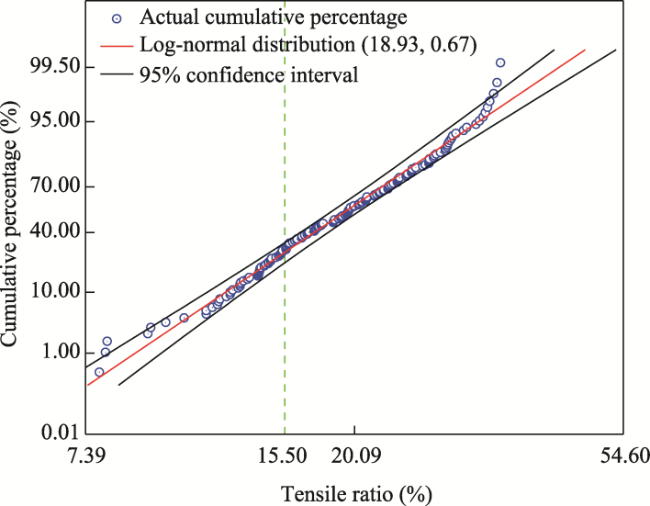
| Log-normal distribution | μ | 0.24 | 6.49 | 146.04 | 18.93 | 775.37 |
| σ | - | 0.53 | 10.42 | 0.67 | 37.00 | |
| R2 | 0.742 | 0.859 | 0.965 | 0.787 | 0.960 |
Note: In the normal distribution and log-normal distribution, μ is the mean value and σ is the variance. In the Weibull distribution, α is the scale parameter, β is the shape parameter, and u is the position parameter. R2 is the goodness of fit. '-' means that the value is less than 0.01. |
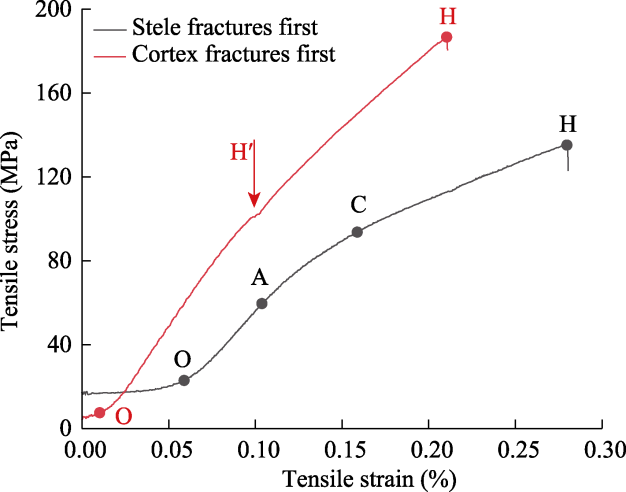
Fig. 6 Tensile stress-strain curves of individual roots from dominant plant species in the alpine meadow layer of the study area. O represents the root begins to straighten and undergo elastic deformation, A represents the start of plastic deformation, C represents the yield point reached, H represents the root fracture, and H′ represents the root cortex fractures. The vertical lines under point H represent the process of rapid stress reduction after root rupture. |
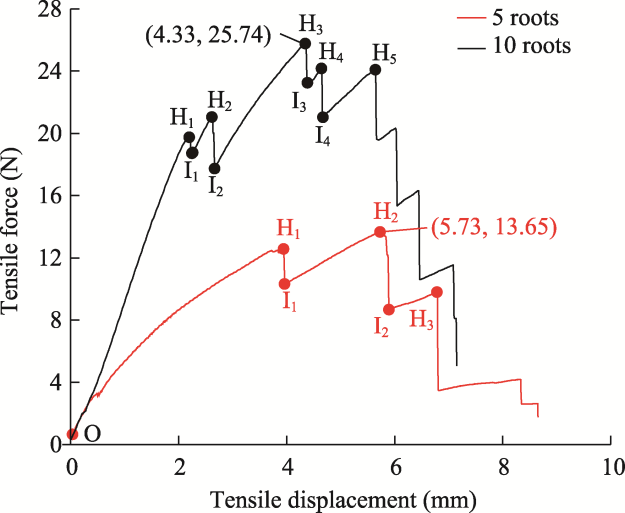
Fig. 7 Typical tensile force-displacement curves from the tensile tests on groups of 5 and 10 roots. Hi, fracture of the ith root in the root group; Ii, condition after the ith root fractures in the root group. The values in the parentheses indicate the coordinate of maximum tensile displacement and force of the root group. |
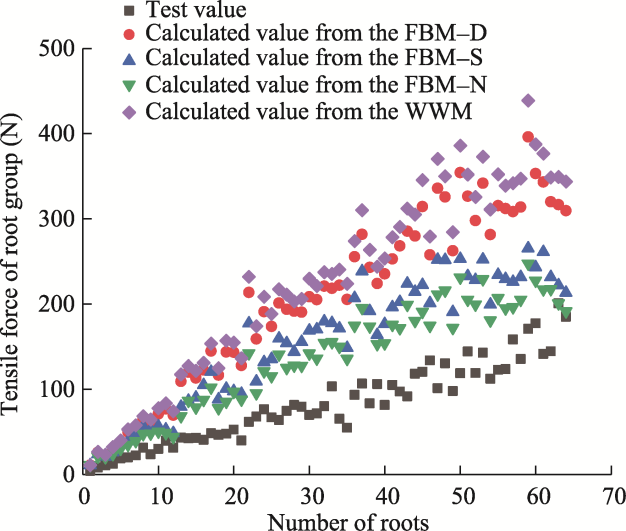
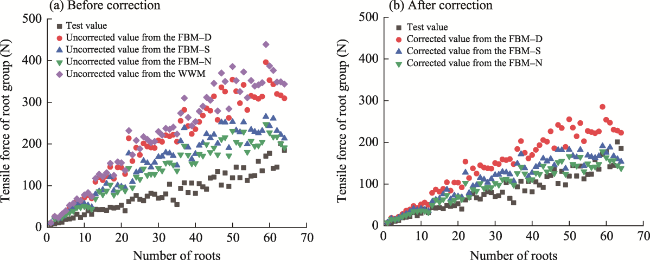
Fig. 8 Comparison between the test tensile force values of root groups and the model calculated tensile force values of root groups obtained from the WWM (Wu-Waldron Model) and three fracture modes in the FBM (Fiber Bundle Model). FBM-D, the tensile force on each single root is proportional to its diameter relative to the total sum of all the root diameters; FBM-S, the cross-sectional stress in the root bundle is uniform; FBM-N, each tensile strength test of individual roots experiences an equal load. |
Table 3 Fitting results of the tensile force values of root groups from tests and three fracture modes in the FBM along with the number of roots |
| Fitting function relationship (y=mx) | Tensile force test | Three fracture modes in the FBM | |||
|---|---|---|---|---|---|
| FBM-D | FBM-S | FBM-N | WWM | ||
| m | 2.54 | 6.11 | 4.55 | 4.01 | 6.68 |
| r | 0.99 | 0.99 | 0.99 | 0.99 | 0.99 |
| R2 | 0.98 | 0.98 | 0.97 | 0.98 | 0.98 |
Note: y is the tensile force; x is the number of roots; m is the fitting function parameter; r is the Person correlation coefficient. FBM-D, the tensile force on each single root is proportional to its diameter relative to the total sum of all the root diameters; FBM-S, the cross-sectional stress in the root bundle is uniform; FBM-N, each tensile strength test of individual roots experiences an equal load. |
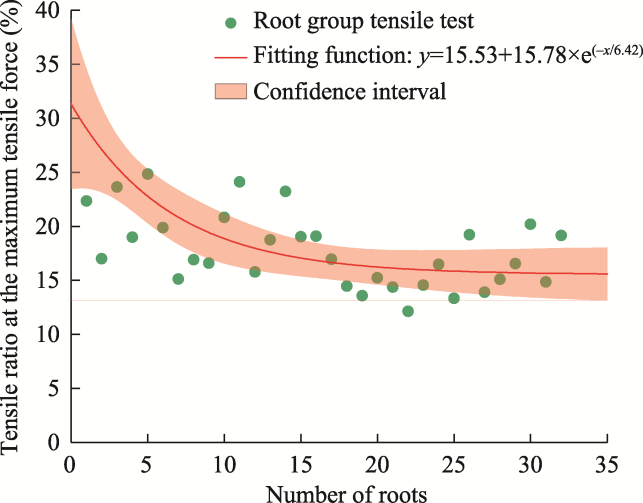
Fig. 9 Relationship between the root group tensile ratio at the maximum tensile force and the number of roots in the root group tensile tests. For the fitted equation, y is the root group tensile ratio, and x is the number of roots. |
Fig. 10 Cumulative curve fitting of log-normal distribution for single root tensile ratio |
Fig. 11 Theoretical tensile force values calculated from the FBM considering tensile displacement correction. (a), before correction; (b), after correction. |
Table 4 Fitting results of the tensile force values of root groups from tests and three fracture modes in the modified FBM along with the number of roots |
| Fitting function relationship (y=mx) | Tensile force test | Three fracture modes in the modified FBM | ||
|---|---|---|---|---|
| FBM-D | FBM-S | FBM-N | ||
| m | 2.54 | 4.40 | 3.27 | 2.88 |
| r | 0.99 | 0.99 | 0.99 | 0.99 |
| R2 | 0.98 | 0.98 | 0.97 | 0.98 |
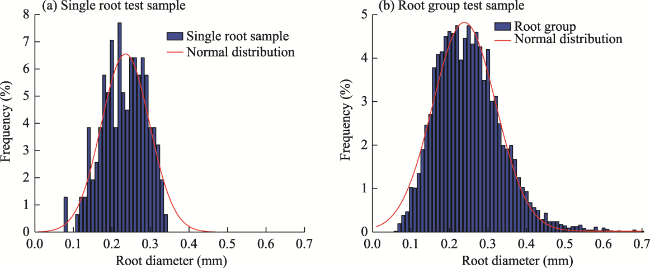
Fig. 12 Statistical comparison of root diameter frequency distributions for single root tensile tests (a) and root group tensile tests (b) |
Table 5 Calculated results of the additional cohesion of the soil provided by the plant roots |
| A (cm2) | Number of roots passing through the shear plane | Theoretical additional cohesion calculated by the model (kPa) | |||
|---|---|---|---|---|---|
| WWM before correction | WWM after correction | FBM-S after correction | FBM-N after correction | ||
| 48 | 81 | 130.84 | 41.43 | 65.38 | 55.78 |
| 86 | 139.02 | 44.02 | 69.40 | 59.22 | |
| 88 | 142.29 | 45.06 | 70.99 | 60.61 | |
| 76 | 122.66 | 38.84 | 61.43 | 52.34 | |
| 144 | 176 | 95.43 | 30.22 | 47.02 | 40.40 |
| 165 | 89.43 | 28.32 | 44.11 | 37.87 | |
| 151 | 81.80 | 25.90 | 40.39 | 34.67 | |
| 172 | 93.25 | 29.53 | 45.96 | 39.48 | |
Note: The empirical correction parameter k′ in the model is 0.38, calculated by dividing the WWM's overestimation ratio. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}