
Benefits and ecological restoration implications of hanging grass fences in Mongolian desert steppe
Received date: 2024-05-19
Revised date: 2024-07-17
Accepted date: 2024-07-23
Online published: 2025-08-12
MIAO Jiamin , LI Shengyu , XU Xinwen , LIU Guojun , WANG Haifeng , FAN Jinglong , Khaulanbek AKHMADI . [J]. Journal of Arid Land, 2024 , 16(11) : 1541 -1561 . DOI: 10.1007/s40333-024-0063-6
Tumbleweeds participate in a common seasonal biological process in temperate grasslands, creating hanging grass fences during the grass-withering season that result in distinct ecological phenomena. In this study, we addressed the urgent need to understand and restore the degraded desert steppe in Central Mongolia, particularly considering the observed vegetation edge effects around hanging grass fences. Using field surveys conducted in 2019 and 2021 in the severely degraded desert steppe of Central Mongolia, we assessed vegetation parameters and soil physical and chemical properties influenced by hanging grass fences and identified the key environmental factors affecting vegetation changes. The results indicate that the edge effects of hanging grass fences led to changes in species distributions, resulting in significant differences in species composition between the desert steppe's interior and edge areas. Vegetation parameters and soil physical and chemical properties exhibited nonlinear responses to the edge effects of hanging grass fences, with changes in vegetation coverage, aboveground biomass, and soil sand content peaking at 26.5, 16.5, and 6.5 m on the leeward side of hanging grass fences, respectively. In the absence of sand dune formation, the accumulation of soil organic carbon and available potassium were identified as crucial factors driving species composition and increasing vegetation coverage. Changes in species composition and plant density were primarily influenced by soil sand content, electrical conductivity, and sand accumulation thickness. These findings suggest that hanging grass fences have the potential to alter vegetation habitats, promote vegetation growth, and control soil erosion in the degraded desert steppe of Central Mongolia. Therefore, in the degraded desert steppe, the restoration potential of hanging grass fences during the enclosure process should be fully considered.
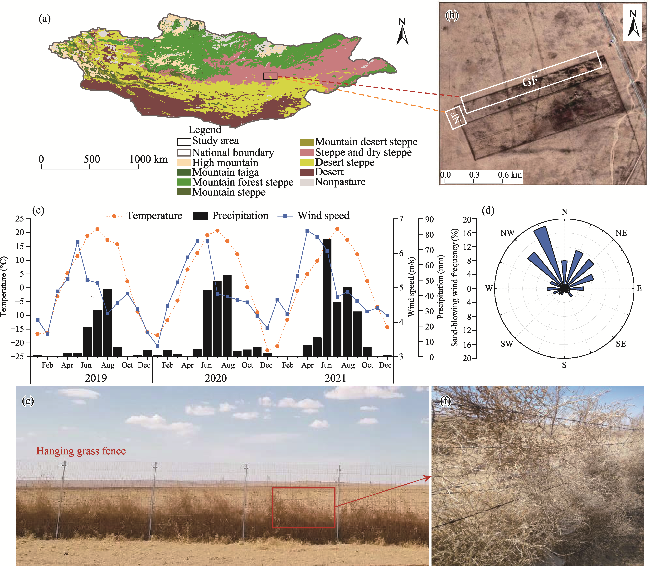
Fig. 1 Overview of the study area, climate characteristics, and hanging grass fences. (a), location of the study area (the study area is located in the ecotone of steppe and desert steppe in Central Mongolia); (b), image showing the experimental design; (c), monthly variations in temperature, precipitation, and wind speed during the sampling period (2019-2021); (d), frequency of sand-driving winds at 10 m in height with wind rose directions; (e), photo showing the hanging grass fence during the grass-withering season (April) in 2019; (f), photo showing the tumbleweeds hanging and accumulating on the fence in April 2019. Note that Figure 1a is based on the standard map (GS(2016)1666) of the Map Service System (http://bzdt.ch.mnr.gov.cn/), and the boundary of the base map has not been modified. The enclosure in Figure 1b is shown in a true-colour image downloaded from Google Earth, including enclosure with hanging grass fences (abbreviated as GF) and enclosure with non-hanging grass fences (abbreviated as NF). N, north; NE, northeast; E, east; SE, southeast; S, south; SW, southwest; W, west; NW, northwest. The abbreviations are the same in the following figures. |
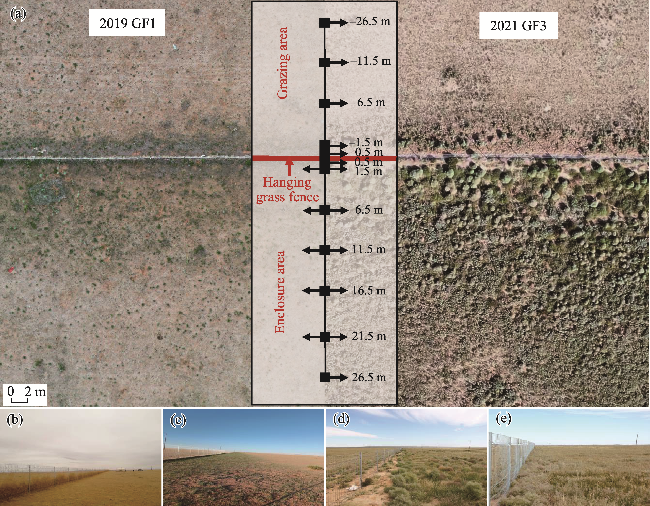
Fig. 2 Experimental design and sampling locations for vegetation and soil survey. (a), drone images showing example locations of transects and sample plots; (b), photo showing vegetation recovery during the early formation of the hanging grass fences in April 2019; (c), photo showing the regreening of the grassland in August 2019 for the first year of enclosure with hanging grass fences (GF1); (d), photo showing the regreening of the grassland in August 2021 for the third year of enclosure with hanging grass fences (GF3); (e), photo showing the third year of enclosure with non-hanging grass fences (NF3). Positive distances indicate sample plots within the enclosure area on the leeward side of the fences, and negative distances indicate sample plots within the grazing area on the windward side of the fences. The abbreviations are the same in the following figures. |
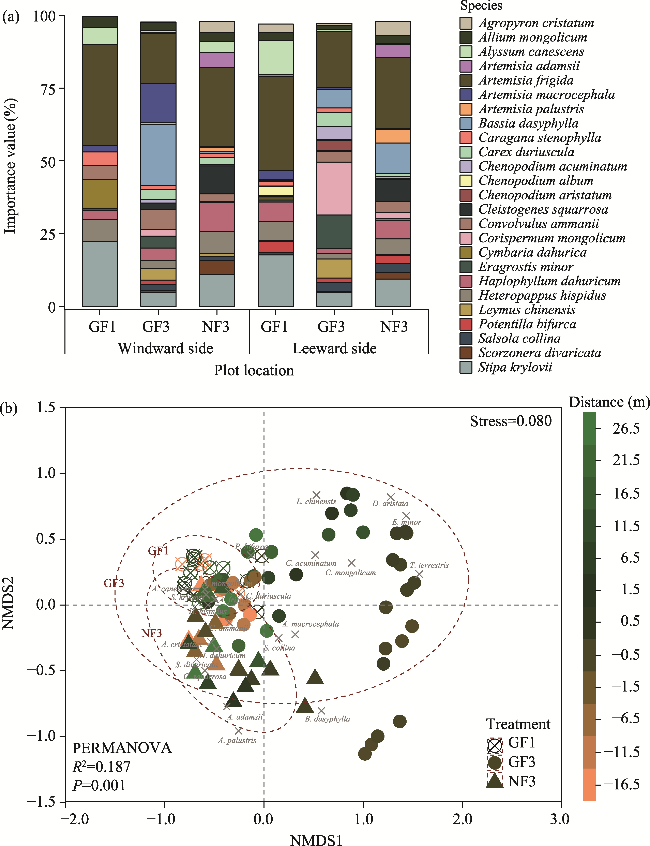
Fig. 3 Responses of species composition to different enclosure treatments (GF1, GF3, and NF3). (a), differences in the importance values of plant species on the windward and leeward sides of fences (the complete data are shown in Tables S1-3); (b), nonmetric multidimensional scaling (NMDS) of species composition, with dispersion ellipses at 95% confidence intervals added. PERMANOVA, permutational multivariate analysis of variance. Ellipse labels represent the 3 enclosure treatments, and the top 20 plant species points in terms of abundance are reflected in the figure. Distance indicates the distance from the sample plots to the fences. |
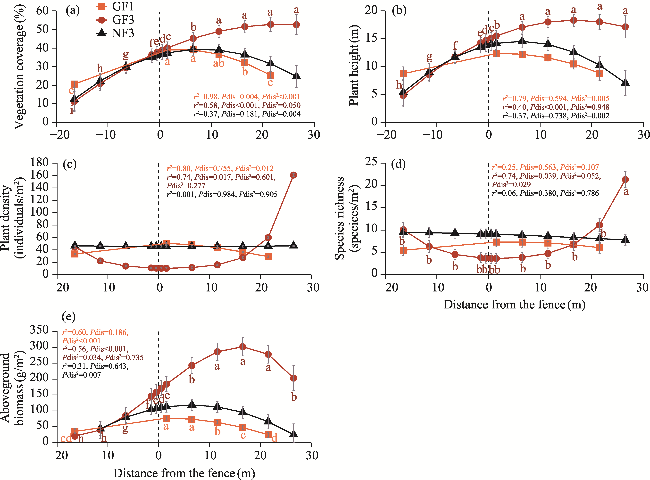
Fig. 4 Comparison of vegetation coverage (a), plant height (b), plant density (c), species richness (d), and aboveground biomass (e) at the interface between the windward and leeward sides of different enclosure treatments. Different lowercase letters indicate significant differences in means (P>0.05). Bars mean standard errors. Positive distances indicate sample plots within the enclosure area on the leeward side of the fences, and negative distances indicate sample plots within the grazing area on the windward side of the fences. Marginal r² refers to the fixed effects only. 'Pdis', 'Pdis2', and 'Ppdis3' represent significance tests for the distance, squared distance, and cubic distance from the fences, respectively. The abbreviations are the same in the following figures. |
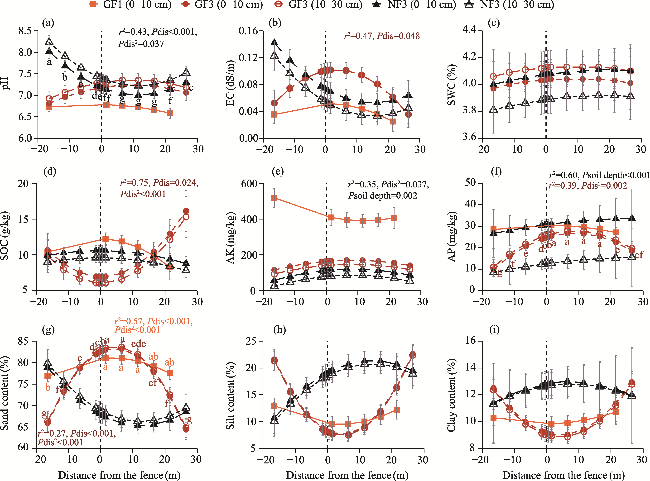
Fig. 5 Comparison of soil physical and chemical properties at the interface between the windward and leeward sides of fences under different enclosure treatments. (a), pH; (b), electrical conductivity (EC); (c), soil water content (SWC); (d), soil organic carbon (SOC) content; (e), available potassium (AK) content; (f), available phosphorus (AP) content; (g), sand content; (h), silt content; (i), clay content. Positive distances indicate sample plots within the enclosure area on the leeward side of the fence, and negative distances indicate sample plots within the grazing area on the windward side of the fence. Different lowercase letters indicate significant differences in means (P>0.05). Bars mean standard errors. Only model r2 values with P<0.05 are labeled in the figure; full model results are provided in Table S4. 'Psoil depth' indicates the significance of soil depth on soil physical and chemical properties. The abbreviations are the same in the following figures. |
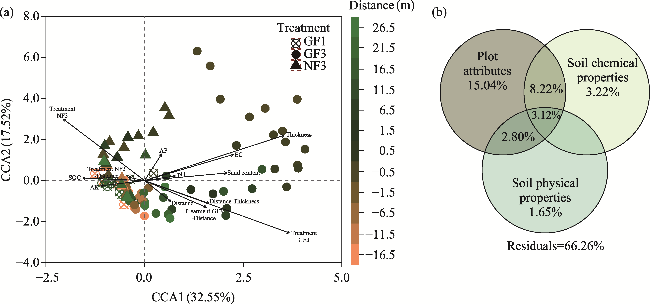
Fig. 6 Impacts of different environmental factors on species composition. (a), canonical correspondence analysis (CCA) of species composition at varying distances from the fence under different enclosure treatments; (b), contributions of different groups of environmental factors to species composition calculated by variation partitioning. Arrows represent the nine significant environmental factors selected for forward selection. "Treatment" represents the three different enclosure treatments: GF1, GF3, and NF3. "Thickness" denotes sand accumulation thickness. "Distance-Thickness" represents the interaction between distance from the fence and sand accumulation thickness. Note that factors with contributions to the total variance less than 0.0 are not shown. |
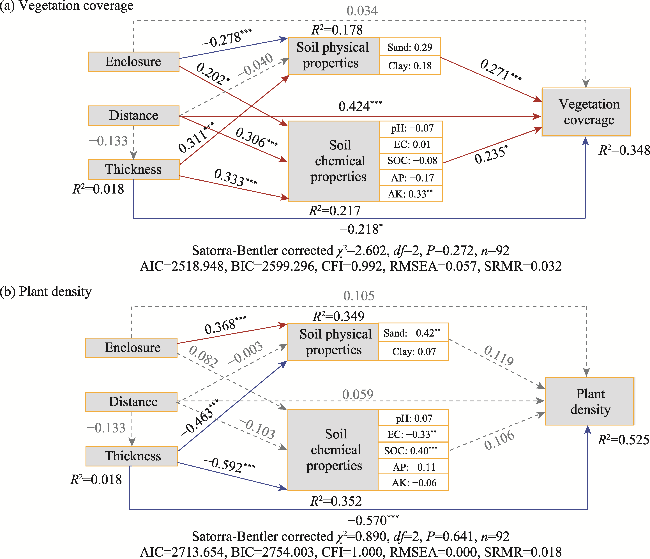
Fig. 7 Impacts of different environmental factors on vegetation coverage (a) and plant density (b) based on structural equation modelling (SEM). Boxes represent the environmental factors included in the model. Arrows indicate the paths, with the numbers on the paths representing standardized regression weights (the thicker the lines, the greater the weights). Solid arrows represent significant paths, with red solid arrows indicating positive effects, blue solid arrows indicating negative effects, and grey dashed arrows indicating non-significant paths. The significances along the paths at P<0.05, P<0.01, and P<0.001 levels are indicated by *, **, and ***, respectively. The standardized estimates and significance are displayed within each soil physical and chemical property box. The total explained variance of the environmental factors (R²) is marked near the box. χ2, chi-square; df, degree of freedom; n, sample size; AIC, Akaike information criterion; BIC, Bayesian information criterion; CFI, comparative fit index; RMSEA, root mean square error of approximation; SRMR, standardized root mean square residual. |
Table S1 Relative importance values of species at different distances from fences in the first year of enclosure with hanging grass fences (GF1) |
| Species | Relative importance value (%) | |||||
|---|---|---|---|---|---|---|
| -16.5 m | 1.5 m | 6.5 m | 11.5 m | 16.5 m | 21.5 m | |
| Bassia dasyphylla | 0.00 | 0.00 | 0.00 | 0.74 | 0.00 | 0.00 |
| Eragrostis minor | 0.90 | 3.48 | 0.00 | 0.00 | 0.00 | 0.00 |
| Chenopodium acuminatum | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Salsola collina | 0.00 | 3.14 | 1.13 | 0.00 | 0.00 | 0.00 |
| Corispermum mongolicum | 0.00 | 1.43 | 0.97 | 0.00 | 0.00 | 0.00 |
| Artemisia frigida | 34.47 | 29.70 | 25.27 | 34.94 | 38.19 | 32.57 |
| Convolvulus ammanii | 5.00 | 0.78 | 0.00 | 0.00 | 1.69 | 0.00 |
| Carex duriuscula | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.27 |
| Chenopodium aristatum | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Allium mongolicum | 3.76 | 0.00 | 3.71 | 3.75 | 0.00 | 5.70 |
| Haplophyllum dahuricum | 2.95 | 7.86 | 6.57 | 4.05 | 7.87 | 7.10 |
| Setaria viridis | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Astragalus sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Chenopodium album | 0.00 | 2.61 | 8.06 | 3.96 | 0.00 | 0.00 |
| Stipa krylovii | 22.32 | 19.99 | 14.85 | 14.79 | 19.10 | 19.74 |
| Alyssum canescens | 5.94 | 15.62 | 4.49 | 13.50 | 13.16 | 11.33 |
| Cleistogenes squarrosa | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Leymus chinensis | 0.00 | 1.86 | 0.00 | 0.00 | 0.00 | 0.00 |
| Sibbaldianthe adpressa | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Tribulus terrestris | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Caragana stenophylla | 4.55 | 2.11 | 0.00 | 3.39 | 0.00 | 2.80 |
| Allium polyrhizum | 1.63 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Artemisia scoparia | 0.00 | 0.00 | 2.36 | 0.00 | 5.70 | 0.00 |
| Heteropappus hispidus | 7.62 | 2.97 | 6.72 | 7.86 | 8.63 | 5.69 |
| Caragana microphylla | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Asparagus dahurica | 0.00 | 2.00 | 3.21 | 0.00 | 0.00 | 0.00 |
| Artemisia macrocephala | 2.30 | 0.00 | 9.81 | 3.51 | 0.00 | 3.15 |
| Oxytropis sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cymbaria dahurica | 9.75 | 0.00 | 0.00 | 0.00 | 0.00 | 3.50 |
| Potentilla bifurca | 0.00 | 0.00 | 2.41 | 7.00 | 2.72 | 7.14 |
| Agropyron cristatum | 0.00 | 7.58 | 7.35 | 0.00 | 0.00 | 0.00 |
| Scorzonera sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Carex sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Artemisia palustris | 0.00 | 0.00 | 1.60 | 0.00 | 0.00 | 0.00 |
| Artemisia adamsii | 0.00 | 0.00 | 1.49 | 2.51 | 0.00 | 0.00 |
Note: Positive distances indicate sample plots within the enclosure area on the leeward side of the fences, and negative distances indicate sample plots within the grazing area on the windward side of the fences. |
Table S2 Relative importance values of species at different distances from fences in the third year of enclosure with hanging grass fences (GF3) |
| Species | Relative importance value (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -16.5 m | -11.5 m | -6.5 m | -1.5 m | -0.5 m | 0.5 m | 1.5 m | 6.5 m | 11.5 m | 16.5 m | 21.5 m | 26.5 m | |
| Bassia dasyphylla | 57.22 | 47.38 | 0.00 | 0.61 | 0.00 | 12.87 | 13.30 | 0.00 | 3.95 | 5.21 | 3.97 | 5.78 |
| Eragrostis minor | 10.30 | 8.87 | 0.36 | 0.00 | 0.00 | 42.85 | 15.23 | 10.71 | 6.93 | 5.97 | 0.00 | 0.00 |
| Chenopodium acuminatum | 0.00 | 0.00 | 3.80 | 2.48 | 0.00 | 7.33 | 5.91 | 1.88 | 4.45 | 4.99 | 1.94 | 3.56 |
| Salsola collina | 0.00 | 6.75 | 2.27 | 0.00 | 1.77 | 6.81 | 5.01 | 0.00 | 2.51 | 2.08 | 2.90 | 1.55 |
| Corispermum mongolicum | 4.75 | 6.90 | 0.00 | 0.00 | 0.00 | 24.81 | 47.39 | 11.56 | 21.05 | 18.70 | 1.52 | 2.39 |
| Artemisia frigida | 1.15 | 0.00 | 29.14 | 31.28 | 25.59 | 0.00 | 0.00 | 19.44 | 21.60 | 26.51 | 31.80 | 36.13 |
| Convolvulus ammanii | 0.00 | 0.00 | 5.81 | 9.78 | 18.16 | 0.00 | 0.00 | 4.17 | 3.48 | 4.85 | 7.87 | 5.00 |
| Carex duriuscula | 0.00 | 0.00 | 7.41 | 6.51 | 2.78 | 0.00 | 0.00 | 5.47 | 3.44 | 4.60 | 11.30 | 8.85 |
| Chenopodium aristatum | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 7.74 | 7.11 | 2.42 | 4.83 | 0.59 | 1.58 | 0.00 |
| Allium mongolicum | 0.00 | 0.00 | 7.72 | 4.01 | 1.49 | 0.00 | 0.00 | 2.17 | 1.35 | 1.02 | 2.40 | 3.65 |
| Haplophyllum dahuricum | 0.00 | 0.00 | 6.81 | 4.16 | 10.12 | 0.00 | 0.00 | 4.60 | 2.13 | 3.69 | 0.83 | 0.00 |
| Setaria viridis | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.41 | 0.00 | 0.00 | 3.73 | 0.00 | 0.00 |
| Astragalus sp. | 0.00 | 0.00 | 0.00 | 3.19 | 4.83 | 0.00 | 0.00 | 0.77 | 0.00 | 0.00 | 0.00 | 3.38 |
| Chenopodium album | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.69 | 0.87 | 0.00 | 0.00 |
| Stipa krylovii | 0.00 | 0.00 | 10.24 | 3.06 | 10.89 | 0.00 | 0.00 | 3.24 | 4.05 | 6.22 | 10.74 | 9.89 |
| Alyssum canescens | 0.00 | 0.00 | 0.69 | 2.47 | 0.00 | 0.00 | 0.00 | 1.17 | 1.01 | 0.00 | 1.66 | 0.00 |
| Cleistogenes squarrosa | 2.15 | 0.00 | 2.36 | 3.78 | 2.83 | 0.00 | 0.00 | 0.00 | 0.00 | 0.48 | 1.93 | 0.38 |
| Leymus chinensis | 0.00 | 0.00 | 3.40 | 9.43 | 7.18 | 0.00 | 0.00 | 13.07 | 15.18 | 2.74 | 6.53 | 8.58 |
| Sibbaldianthe adpressa | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.54 | 1.39 | 2.23 |
| Tribulus terrestris | 0.00 | 6.26 | 0.00 | 0.00 | 0.00 | 1.47 | 3.64 | 0.62 | 0.89 | 0.00 | 0.00 | 0.74 |
| Caragana stenophylla | 0.00 | 0.00 | 2.10 | 1.69 | 2.97 | 0.00 | 0.00 | 1.82 | 0.00 | 7.55 | 0.00 | 2.02 |
| Allium polyrhizum | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.44 | 0.99 | 1.55 |
| Artemisia scoparia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.81 | 0.00 |
| Heteropappus hispidus | 0.00 | 0.00 | 1.24 | 7.51 | 5.04 | 0.00 | 0.00 | 8.29 | 0.00 | 0.00 | 1.00 | 3.55 |
| Caragana microphylla | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.70 | 0.00 |
| Asparagus dahurica | 0.00 | 0.00 | 1.46 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.46 | 0.00 | 1.39 |
| Artemisia macrocephala | 22.81 | 23.84 | 7.97 | 7.60 | 4.30 | 0.00 | 0.00 | 3.91 | 0.00 | 0.00 | 0.00 | 0.00 |
| Oxytropis sp. | 0.00 | 0.00 | 1.05 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cymbaria dahurica | 0.00 | 0.00 | 1.29 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Potentilla bifurca | 0.00 | 0.00 | 5.77 | 1.41 | 0.00 | 0.00 | 0.00 | 2.94 | 2.30 | 1.03 | 4.15 | 0.75 |
| Agropyron cristatum | 0.00 | 0.00 | 0.00 | 0.00 | 1.34 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4.70 | 0.00 |
| Scorzonera sp. | 0.00 | 0.00 | 2.06 | 1.02 | 0.00 | 0.00 | 0.00 | 0.89 | 0.00 | 0.00 | 0.00 | 0.92 |
| Carex sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.72 | 0.00 | 0.00 | 0.85 | 0.00 | 0.00 | 0.00 | 0.00 |
| Artemisia palustris | 0.00 | 0.00 | 3.17 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Artemisia adamsii | 1.62 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
Note: Positive distances indicate sample plots within the enclosure area on the leeward side of the fences, and negative distances indicate sample plots within the grazing area on the windward side of the fences. |
Table S3 Relative importance values of species at different distances from fences in the third year of enclosure with non-hanging grass fences (NF3) |
| Species | Relative importance value (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| -16.5 m | -11.5 m | -6.5 m | -1.5 m | -0.5 m | 0.5 m | 1.5 m | 6.5 m | 11.5 m | 16.5 m | 21.5 m | 26.5 m | |
| Bassia dasyphylla | 3.47 | 0.00 | 0.00 | 0.00 | 0.00 | 40.88 | 17.53 | 6.66 | 0.00 | 8.25 | 0.00 | 0.00 |
| Eragrostis minor | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Chenopodium acuminatum | 2.76 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.96 | 0.00 | 0.00 | 2.21 | 0.00 | 0.00 |
| Salsola collina | 2.99 | 0.00 | 0.00 | 1.69 | 1.70 | 2.96 | 5.80 | 9.26 | 0.00 | 1.09 | 1.40 | 0.00 |
| Corispermum mongolicum | 2.02 | 0.00 | 0.00 | 0.00 | 0.00 | 10.97 | 3.99 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Artemisia frigida | 18.03 | 30.11 | 25.68 | 25.11 | 36.72 | 4.37 | 22.05 | 20.85 | 23.27 | 40.23 | 36.99 | 22.71 |
| Convolvulus ammanii | 0.00 | 0.00 | 7.63 | 5.24 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 10.79 | 14.09 |
| Carex duriuscula | 0.00 | 0.00 | 5.22 | 6.09 | 0.00 | 0.00 | 0.00 | 1.24 | 1.53 | 1.94 | 0.00 | 2.20 |
| Chenopodium aristatum | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Allium mongolicum | 1.78 | 3.53 | 4.54 | 0.00 | 2.42 | 0.00 | 0.00 | 0.00 | 1.92 | 11.66 | 3.99 | 2.11 |
| Haplophyllum dahuricum | 9.69 | 6.98 | 11.60 | 13.18 | 7.53 | 10.46 | 10.01 | 1.73 | 10.28 | 3.18 | 0.00 | 9.49 |
| Setaria viridis | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.74 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Astragalus sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.22 | 2.38 | 0.91 | 0.00 | 0.00 | 2.34 |
| Chenopodium album | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.37 | 1.69 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Stipa krylovii | 7.40 | 19.59 | 12.17 | 6.26 | 9.81 | 7.10 | 8.96 | 12.42 | 11.10 | 2.01 | 12.75 | 11.69 |
| Alyssum canescens | 1.52 | 6.05 | 2.46 | 3.04 | 5.97 | 0.84 | 0.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 |
| Cleistogenes squarrosa | 7.07 | 8.21 | 16.19 | 11.00 | 5.68 | 0.00 | 0.00 | 9.10 | 16.70 | 4.02 | 8.61 | 15.09 |
| Leymus chinensis | 0.00 | 0.00 | 0.00 | 0.00 | 5.81 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.78 | 0.00 |
| Sibbaldianthe adpressa | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Tribulus terrestris | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Caragana stenophylla | 3.07 | 4.28 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Allium polyrhizum | 0.00 | 0.00 | 0.00 | 2.52 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4.27 |
| Artemisia scoparia | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Heteropappus hispidus | 7.06 | 4.55 | 3.75 | 11.05 | 11.66 | 2.14 | 3.69 | 0.00 | 9.65 | 8.89 | 5.51 | 7.39 |
| Caragana microphylla | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Asparagus dahurica | 0.00 | 2.86 | 3.69 | 0.00 | 5.17 | 0.00 | 0.00 | 0.00 | 0.00 | 5.21 | 0.00 | 0.00 |
| Artemisia macrocephala | 2.92 | 0.00 | 0.00 | 0.00 | 0.00 | 2.38 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Oxytropis sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cymbaria dahurica | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4.50 |
| Potentilla bifurca | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 8.62 | 11.91 | 0.00 |
| Agropyron cristatum | 5.45 | 1.68 | 2.33 | 6.58 | 1.91 | 4.43 | 14.73 | 2.51 | 6.79 | 2.69 | 3.00 | 0.00 |
| Scorzonera sp. | 1.87 | 3.24 | 4.72 | 8.24 | 5.64 | 0.00 | 0.00 | 0.00 | 8.71 | 0.00 | 3.27 | 4.11 |
| Carex sp. | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Artemisia palustris | 6.02 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 10.29 | 22.63 | 0.00 | 0.00 | 0.00 | 0.00 |
| Artemisia adamsii | 16.89 | 8.95 | 0.00 | 0.00 | 0.00 | 10.37 | 1.55 | 11.21 | 7.82 | 0.00 | 0.00 | 0.00 |
Note: Positive distances indicate sample plots within the enclosure area on the leeward side of the fences, and negative distances indicate sample plots within the grazing area on the windward side of the fences. |
Table S4 Effects of the duration of enclosure with hanging grass fences (GF) and the presence of hanging grasses on vegetation parameters and soil physical and chemical properties based on the linear mixed model (LMM) |
| Variable | Goodness-of-fit | Intercept | Distance from the fence (m) | DiffGF1-GF3 | SAT | Distance-Year 2021 | Soil depth (0-10 cm) | |
|---|---|---|---|---|---|---|---|---|
| Marginal r² | Conditional r² | |||||||
| GF1 vs. GF3 | ||||||||
| Vegetation coverage | 0.525 | 0.585 | 31.397*** (3.563) | 0.241 (0.186) | 8.150* (3.684) | 0.096 (0.155) | 0.798** (0.241) | - |
| Plant height | 0.460 | 0.546 | 10.867*** (1.342) | 0.037 (0.007) | 2.324 (1.322) | 0.067 (0.056) | 0.295** (0.086) | - |
| Aboveground biomass | 0.629 | 0.670 | 55.865* (23.022) | -0.049 (1.248) | 109.238*** (24.609) | 2.075 (1.041) | 6.862*** (1.615) | - |
| Species richness | 0.557 | 0.560 | 6.495*** (1.052) | 1.004 (1.003) | 1.408*** (1.007) | 0.980*** (1.003) | 0.996 (1.004) | - |
| Plant density | 0.904 | 0.904 | 46.390*** (1.058) | 1.001 (1.006) | 0.898 (1.094) | 0.975*** (1.002) | 0.999 (1.008) | - |
| pH | 0.414 | 0.586 | 6.617*** (0.126) | -0.001 (0.005) | 0.490*** (0.101) | 0.007 (0.004) | 0.011 (0.007) | - |
| SOC | 0.256 | 0.320 | 10.220*** (0.580) | 0.007 (0.030) | -0.413 (0.650) | -0.048 (0.030) | -0.101* (0.043) | - |
| EC | 0.459 | 0.529 | 0.038** (0.008) | 0.000 (0.000) | 0.021* (0.009) | 0.002*** (0.000) | 0.001 (0.001) | - |
| AP | 0.656 | 0.913 | 32.226*** (2.647) | -0.054 (0.166) | -13.076*** (2.482) | 0.178** (0.060) | 0.421 (0.216) | - |
| AK | 0.955 | 0.972 | 434.036*** (17.475) | -3.688* (1.613) | -295.078*** (23.429) | 0.583 (0.576) | 3.920 (2.101) | - |
| Sand content | 0.344 | 0.356 | 77.912*** (1.492) | 0.032 (0.099) | -4.051* (1.904) | 0.363*** (0.082) | 0.163 (0.128) | - |
| Silt content | 0.336 | 0.350 | 11.714*** (1.234) | -0.042 (0.081) | 3.581* (1.568) | -0.290*** (0.068) | -0.114 (0.105) | - |
| Clay content | 0.363 | 0.669 | 10.096*** (0.588) | 0.012 (0.050) | -0.252 (0.737) | -0.052** (0.018) | -0.053 (0.066) | - |
| Vegetation coverage | 0.369 | 0.389 | 32.035*** (3.293) | 0.247 (0.220) | 4.982 (3.968) | -0.125 (0.093) | 0.654* (0.271) | - |
| Plant height | 0.315 | 0.325 | 11.714*** (1.171) | 0.023 (0.083) | 0.133 (1.472) | 0.093* (0.035) | 0.285** (0.103) | - |
| Aboveground biomass | 0.383 | 0.471 | 102.824** (26.800) | 0.472 (1.377) | 42.495 (26.267) | 0.411 (0.587) | 5.777** (1.698) | - |
| Species richness | 0.638 | 0.672 | 8.933*** (1.081) | 0.995 (1.004) | 1.015 (1.086) | 0.980*** (1.002) | 1.006 (1.005) | - |
| Plant density | 0.666 | 0.674 | 46.109*** (1.100) | 1.000 (1.007) | 0.905 (1.124) | 0.971*** (1.003) | 1.002 (1.008) | - |
| pH | 0.114 | 0.288 | 7.386*** (0.127) | -0.015** (0.005) | -0.211* (0.091) | 0.003 (0.002) | 0.022*** (0.006) | -0.148* (0.067) |
| SOC | 0.215 | 0.219 | 10.038*** (0.492) | -0.028 (0.032) | 0.636 (0.557) | -0.080*** (0.013) | 0.033 (0.039) | -0.408 (0.457) |
| EC | 0.259 | 0.299 | 0.078*** (0.012) | -0.002** (0.001) | -0.025* (0.012) | 0.002*** (0.000) | 0.002* (0.009) | 0.006 (0.001) |
| AP | 0.308 | 0.512 | 16.987*** (3.454) | 0.166 (0.370) | -2.288 (3.931) | 0.273*** (0.065) | 0.276 (0.454) | 4.070* (1.909) |
| AK | 0.366 | 0.382 | 69.350*** (9.090) | 0.573 (0.521) | 44.403*** (9.564) | 0.903*** (0.221) | 0.232 (0.643) | 26.387*** (7.516) |
| Sand content | 0.489 | 0.493 | 73.785*** (1.238) | -0.179 (0.232) | 5.364** (2.002) | 0.145*** (0.037) | 0.260 (0.361) | -2.150 (1.195) |
| Silt content | 0.425 | 0.435 | 13.740*** (0.990) | 0.164 (0.180) | -3.015 (1.587) | -0.105*** (0.029) | −0.208 (0.221) | 1.741 (0.927) |
| Clay content | 0.486 | 0.486 | 12.520*** (0.408) | 0.015 (0.079) | -2.444*** (0.664) | -0.039** (0.012) | -0.052 (0.097) | 0.409 (0.407) |
| SWC | 0.004 | 0.059 | 4.007 (0.125) | 0.002 (0.006) | 0.039 (0.121) | 0.001 (0.003) | -0.001 (0.008) | 0.002 (0.093) |
Note: The goodness of fit, intercept, coefficient, and standard error (value in the parenthese) of LMM are presented. DiffGF1-GF3, the difference between GF3 and GF1; DiffGF1-NF3, the difference between GF3 and NF3; SAT, sand accumulation thickness; Distance-Year 2021, interaction effects with fence distance and year. EC, electrical conductivity; SWC, soil water content; SOC, soil organic carbon; AK, available potassium; AP, available phosphorus. "-" indicates that the variable is not included in the model calculation. *, **, and *** mean coefficients outside the 95.0%, 99.0% and 99.9% confidence intervals, respectively. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}