
Diversity and plant growth-promoting properties of culturable bacteria associated with three halophytes in an arid land, Northwest China
Received date: 2024-09-27
Revised date: 2025-02-20
Accepted date: 2025-03-24
Online published: 2025-08-12
HUANG Yin , ZHANG Xiaoye , MA Jinbiao , JIAO Haocheng , Murad MUHAMMAD , Rashidin ABDUGHENI , Vyacheslav SHURIGIN , Dilfuza EGAMBERDIEVA , LI Li . [J]. Journal of Arid Land, 2025 , 17(5) : 696 -713 . DOI: 10.1007/s40333-025-0015-9
Salt-tolerant bacteria associated with halophytes enhance plant resistance and adaptation to environmental stress. The purpose of this study was to investigate the diversity and plant-beneficial traits of bacteria associated with three halophytes in an arid land, Northwest China. The bacterial strains were isolated from the roots, shoots, rhizosphere, and bulk soil of three halophytes, i.e., Salicornia europaea L., Kalidium foliatum (Pall.) Moq., and Suaeda aralocaspica (Bunge) Freitag & Schütze, collected from the saline soils near to the Wujiaqu City, Xinjiang, Northwest China. A total of 567 strains were isolated and identified from these three halophytes belonging to 4 phyla, 6 classes, 25 orders, 36 families, and 66 genera, including 147 potential novel species. A total of 213 strains exhibited one or more plant growth- promoting properties, while 20 strains demonstrated multiple in vitro plant growth-promoting activities, including phosphate solubilization, nitrogen fixation, siderophore production, and production of hydrolytic enzymes such as protease and cellulase. Our findings showed that halophytes in the arid land harbor diverse bacteria with the potential to enhance plant growth and adaptability under challenging environmental conditions.
Key words: halophytes; endophytic bacteria; rhizosphere bacteria; diversity; functional strains
Table 1 Sample information and abbreviation from the three halophytes |
| Sample | Salicornia europaea L. (P1) | Kalidium foliatum (Pall.) Moq. (P2) | Suaeda aralocaspica (Bunge) Freitag & Schütze (P3) |
|---|---|---|---|
| Root and shoot (PE) | P1PE | P2PE | P3PE |
| Rhizosphere soil (RR) | P1RR | P2RR | P3RR |
| Bulk soil (RS) | P1RS | P2RS | P3RS |
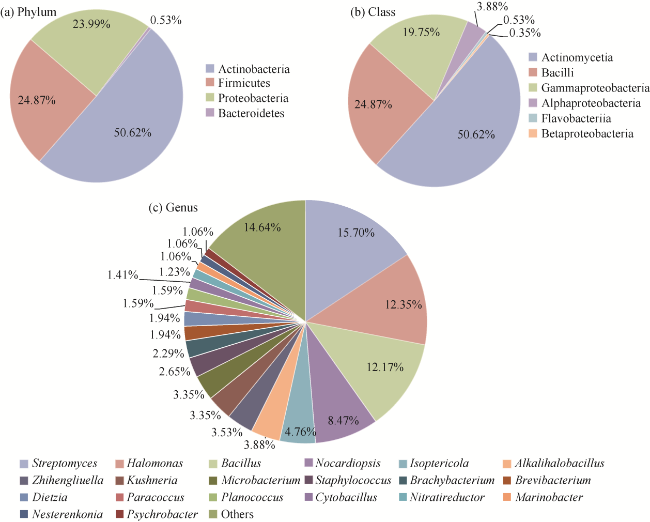
Fig. 1 Community composition of culturable bacteria from the three halophytes at phylum (a), class (b), and genus (c) (top 20 dominant genera) levels |
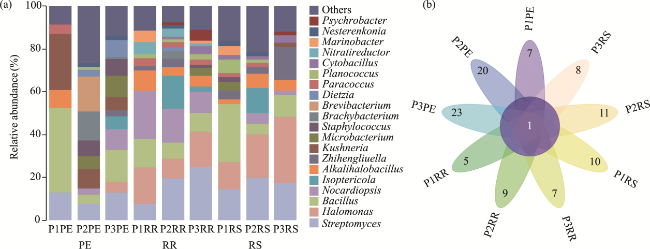
Fig. 2 Community composition at the genus level (top 20 dominant genera) (a) and flower diagram (b) at the species level of culturable bacteria isolated from different samples of the three halophytes (P1, P2, and P3). PE, RR, and RS indicate samples form root and shoot, rhizosphere soil, and bulk soil, respecitively. The detailed sample information is shown in Table 1. The abbreviations are the same in the following tables and figures. |
Table 2 Taxa and diversity of culturable bacteria |
| Group | Index | P1PE | P2PE | P3PE | P1RR | P2RR | P3RR | P1RS | P2RS | P3RS |
|---|---|---|---|---|---|---|---|---|---|---|
| Taxa | Phylum | 3 | 3 | 3 | 3 | 3 | 4 | 3 | 4 | 3 |
| Class | 4 | 5 | 4 | 4 | 4 | 5 | 4 | 5 | 3 | |
| Order | 6 | 11 | 16 | 11 | 15 | 13 | 9 | 12 | 11 | |
| Family | 6 | 16 | 18 | 13 | 17 | 15 | 11 | 14 | 12 | |
| Genus | 7 | 24 | 25 | 16 | 21 | 20 | 18 | 22 | 16 | |
| Isolates | 23 | 67 | 101 | 53 | 77 | 80 | 48 | 60 | 58 | |
| Diversity | Richness | 13.00 | 36.00 | 47.00 | 28.00 | 42.00 | 37.00 | 27.00 | 35.00 | 27.00 |
| Shannon | 2.36 | 3.39 | 3.57 | 3.02 | 3.47 | 3.40 | 3.04 | 3.29 | 2.98 | |
| Simpson | 0.88 | 0.96 | 0.96 | 0.93 | 0.96 | 0.96 | 0.94 | 0.95 | 0.93 | |
| Pielou | 0.92 | 0.93 | 0.93 | 0.91 | 0.93 | 0.94 | 0.92 | 0.92 | 0.90 |
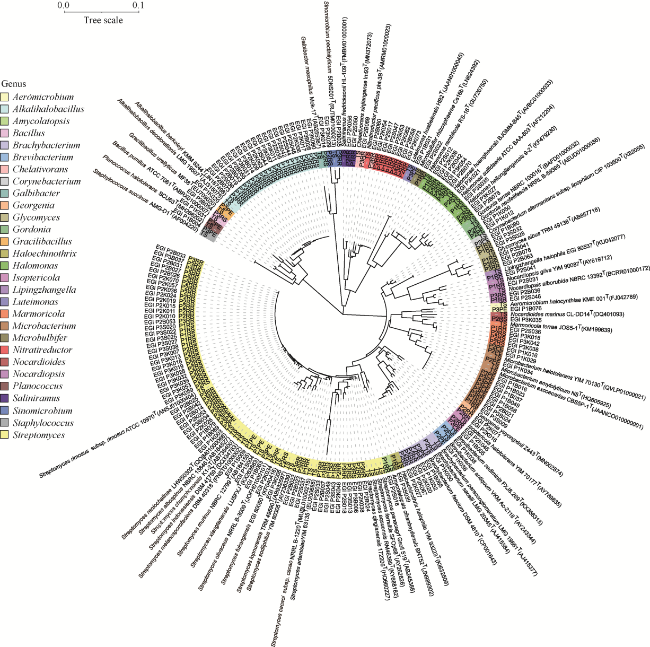
Fig. 3 Phylogenetic tree of potential novel bacterial strains. The effective sequence length was 538 bp. Bar, the number of substitutions per sample. |
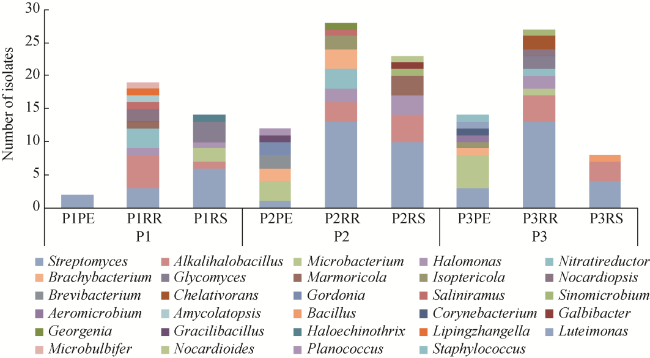
Fig. 4 Diversity of potentially novel bacterial strains isolated at the genus level |
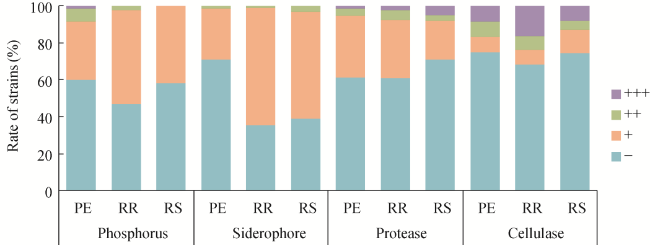
Fig. 5 Screening results in bacterial strains solubilizing phosphate, producing siderophores, protease, and cellulase. ''‒'' indicates negative solubilization ability (E=1); ''+'' indicates weak solubilization ability (1<E≤2); ''++'' indicates moderate solubilization ability (2<E≤3); and ''+++'' indicates strong solubilization ability (E>3). E is the ability to solubilize phosphate and produce siderophores, protease, and cellulose. |
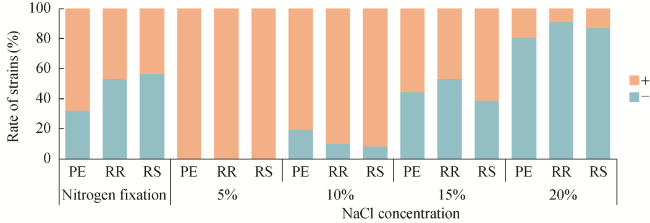
Fig. 6 Ability of nitrogen fixation and salt tolerance in bacterial strains. ''‒'' indicates negative and ''+'' indicates positive for nitrogen fixation and salt tolerance ability. |
Table S1 Unique species of culturable bacteria in different sites |
| Sampling site | Unique species of culturable bacteria |
|---|---|
| P1PE | Alkalihalobacillus pseudofirmus, Bacillus atrophaeus, Corynebacterium mucifaciens, Kocuria polaris, Kushneria marisflavi, Streptomyces murinus, and Streptomyces xiangtanensis |
| P2PE | Advenella kashmirensis subsp. methylica, Arthrobacter gandavensis, Brevibacterium anseongense, Brevibacterium antiquum, Brevibacterium epidermidis, Brevibacterium sediminis, Citricoccus alkalitolerans, Corynebacterium glyciniphilum, Frigoribacterium endophyticum, Gordonia neofelifaecis, Gordonia terrae, Gracilibacillus ureilyticus, Kushneria pakistanensis, Mammaliicoccus sciuri, Microbacterium hydrocarbonoxydans, Mycolicibacterium frederiksbergense, Oceanobacillus picturae, Pelagibacterium luteolum, Salinicola corii, and Streptomyces melanosporofaciens |
| P3PE | Aeromicrobium halocynthiae, Corynebacterium afermentans subsp. lipophilum, Demequina activiva, Demequina aestuarii, Halomonas titanicae, Luteimonas huabeiensis, Lysobacter spongiicola, Microbacterium amylolyticum, Microbacterium pumilum, Nesterenkonia halobia, Nocardiopsis alba, Ornithinimicrobium pekingense, Priestia filamentosa, Promicromonospora xylanilytica, Pseudarthrobacter oxydans, Rhizobium marinum subsp. pelagicum, Staphylococcus cohnii, Staphylococcus saprophyticus subsp. bovis, Streptomyces albidoflavus, Streptomyces daqingensis, Streptomyces microflavus, Streptomyces pseudovenezuelae, and Streptomyces violascens |
| P1RR | Halobacillus litoralis, Lipingzhangella halophila, Paracoccus marcusii, Planococcus plakortidis, and Streptomyces sodiiphilus |
| P2RR | Georgenia yuyongxinii, Halomonas stenophila, Marinococcus salis, Prauserella aidingensis, Streptomyces anulatus, Streptomyces diacarni, Streptomyces indonesiensis, Streptomyces rimosus subsp. rimosus, and Zhihengliuella salsuginis |
| P3RR | Anaerobacillus isosaccharinicus, Chelativorans xinjiangense, Halomonas olivaria, Microbacterium suaedae, Nocardiopsis chromatogenes, Planococcus salinarum, and Streptomyces monticola |
| P1RS | Haloechinothrix halophila, Halomonas montanilacus, Halomonas sulfidaeris, Marinobacter lipolyticus, Nesterenkonia aurantiaca, Paenisporosarcina quisquiliarum, Planococcus antarcticus, Planomicrobium iranicum, Streptomyces albospinus, and Virgibacillus salarius |
| P2RS | Arthrobacter ruber, Exiguobacterium mexicanum, Galbibacter mesophilus, Halomonas huangheensis, Isoptericola salitolerans, Myceligenerans xiligouense, Nocardioides marinus, Streptomyces aqsuensis, Streptomyces artemisiae, Streptomyces flocculus, and Streptomyces lopnurensis |
| P3RS | Alcanivorax xenomutans, Halomonas elongata, Halomonas ventosae, Salininema proteolyticum, Streptomyces cellulosae, Streptomyces flavovirens, Streptomyces lusitanus, and Streptomyces panacagri |
Table S2 Potential plant-beneficial traits of bacteria associated with the three halophytes |
| Strain | Phosphorus | Siderophore | Hydrolytic enzyme | Nitrogen fixation | Salt-tolerant concentration | ||||
|---|---|---|---|---|---|---|---|---|---|
| Protease | Cellulase | 5.00% | 10.00% | 15.00% | 20.00% | ||||
| EGI P1B004 | + | + | ‒ | ‒ | + | + | + | + | + |
| EGI P1B007 | + | ‒ | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P1B018 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P1B021 | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ | ‒ |
| EGI P1B030 | + | + | ‒ | ‒ | + | + | ‒ | ‒ | ‒ |
| EGI P1B031 | ‒ | ‒ | ‒ | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P1B032 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P1B037 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P1B041 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P1B044 | ‒ | ‒ | +++ | +++ | + | + | + | + | ‒ |
| EGI P1B047 | ‒ | ‒ | ‒ | + | ‒ | + | ‒ | ‒ | ‒ |
| EGI P1B048 | + | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P1B049 | ‒ | ‒ | + | ‒ | + | + | + | + | ‒ |
| EGI P1B050 | ‒ | ‒ | ‒ | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P1B053 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P1B057 | ++ | ‒ | + | ++ | + | + | + | + | ‒ |
| EGI P1B059 | + | ‒ | + | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P1B060 | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P1B065 | ‒ | ‒ | + | ‒ | + | + | + | + | ‒ |
| EGI P1B067 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P1B068 | ‒ | ‒ | + | ++ | + | + | + | ‒ | ‒ |
| EGI P1B072 | ‒ | + | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P1B073 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | + |
| EGI P1B074 | ‒ | ‒ | + | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P1B076 | ‒ | ‒ | ‒ | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P1B078 | + | + | + | ‒ | + | + | + | ‒ | ‒ |
| EGI P1B079 | ‒ | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P1B081 | ++ | + | + | + | + | + | + | + | ‒ |
| EGI P1B088 | + | + | ++ | +++ | + | + | ‒ | ‒ | ‒ |
| EGI P1B089 | + | + | ++ | +++ | + | + | ‒ | ‒ | ‒ |
| EGI P1B090 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P1B091 | + | ‒ | + | ++ | + | + | + | ‒ | ‒ |
| EGI P1B094 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P1B101 | ‒ | ‒ | + | ‒ | ‒ | + | + | + | ‒ |
| EGI P1K001 | + | ‒ | ‒ | ‒ | + | + | + | + | + |
| EGI P1K005 | ++ | + | ‒ | ‒ | + | + | + | + | + |
| EGI P1K006 | + | ‒ | + | ++ | + | + | + | + | ‒ |
| EGI P1K020 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P1K021 | ‒ | ‒ | ‒ | +++ | + | + | + | ‒ | ‒ |
| EGI P1K024 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P1K025 | ‒ | ‒ | + | ‒ | + | + | + | + | ‒ |
| EGI P1K026 | ‒ | ‒ | + | ‒ | + | + | + | + | ‒ |
| EGI P1K028 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P1K029 | + | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P1K030 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P1K031 | ‒ | ‒ | ‒ | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P1K036 | ‒ | ‒ | + | ‒ | + | + | + | + | ‒ |
| EGI P1K037 | + | ‒ | + | ‒ | + | + | + | + | ‒ |
| EGI P1K039 | ‒ | ‒ | ‒ | + | ‒ | + | + | ‒ | ‒ |
| EGI P1K041 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P1K042 | + | ‒ | + | ‒ | + | + | + | + | ‒ |
| EGI P1K043 | + | ‒ | + | + | ‒ | + | ‒ | ‒ | ‒ |
| EGI P1K047 | +++ | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P1K050 | ++ | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P1K052 | + | + | + | + | + | + | + | + | ‒ |
| EGI P1K056 | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P1K057 | + | ‒ | ++ | ++ | + | + | + | ‒ | ‒ |
| EGI P1K058 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P1K061 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P1K065 | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ | ‒ |
| EGI P1K067 | ‒ | + | ‒ | ‒ | + | + | ‒ | ‒ | ‒ |
| EGI P1S002 | + | ‒ | + | +++ | + | + | + | + | ‒ |
| EGI P1S006 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P1S007 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P1S009 | + | ‒ | + | ++ | + | + | ‒ | ‒ | ‒ |
| EGI P1S012 | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P1S013 | ++ | ++ | + | ‒ | + | + | + | + | ‒ |
| EGI P1S017 | + | ‒ | + | +++ | + | + | + | ‒ | ‒ |
| EGI P1S018 | + | ‒ | + | ‒ | + | + | + | + | + |
| EGI P1S019 | + | + | + | + | + | + | + | + | ‒ |
| EGI P1S022 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | + |
| EGI P1S023 | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P2B001 | + | + | + | ++ | + | + | + | + | ‒ |
| EGI P2B006 | + | + | ‒ | ‒ | + | + | + | + | + |
| EGI P2B012 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P2B013 | + | + | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P2B016 | ‒ | ‒ | + | ‒ | + | + | + | ‒ | ‒ |
| EGI P2B027 | + | + | ‒ | +++ | + | + | + | ‒ | ‒ |
| EGI P2B029 | + | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2B034 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2B035 | ‒ | ‒ | + | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P2B038 | + | + | + | + | + | + | + | ‒ | ‒ |
| EGI P2B040 | ‒ | + | ‒ | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P2B042 | + | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2B045 | + | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2B046 | + | ‒ | ‒ | + | ‒ | + | + | + | + |
| EGI P2B047 | + | + | +++ | +++ | + | + | + | ‒ | ‒ |
| EGI P2B049 | + | ‒ | + | ++ | + | + | + | ‒ | ‒ |
| EGI P2B050 | + | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2B054 | ++ | + | + | +++ | + | + | + | ‒ | ‒ |
| EGI P2B055 | ‒ | ‒ | ++ | ‒ | + | + | + | + | ‒ |
| EGI P2B056 | + | + | + | ++ | ‒ | + | + | + | ‒ |
| EGI P2B057 | + | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2B058 | + | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2B059 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2B075 | + | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2B077 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2B079 | + | ‒ | ‒ | ‒ | + | + | + | + | + |
| EGI P2K001 | + | + | + | + | + | + | + | + | ‒ |
| EGI P2K007 | + | + | ++ | +++ | + | + | ‒ | ‒ | ‒ |
| EGI P2K008 | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K009 | ‒ | + | + | ‒ | ‒ | + | + | + | ‒ |
| EGI P2K011 | + | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K012 | + | ‒ | +++ | + | ‒ | + | + | ‒ | ‒ |
| EGI P2K017 | + | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2K019 | + | ‒ | + | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2K020 | ‒ | + | + | ‒ | ‒ | + | + | + | ‒ |
| EGI P2K022 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2K028 | + | ‒ | + | +++ | + | + | + | + | ‒ |
| EGI P2K030 | + | ‒ | ‒ | +++ | ‒ | + | + | ‒ | ‒ |
| EGI P2K031 | + | ‒ | + | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K032 | ++ | + | ‒ | + | + | + | + | ‒ | ‒ |
| EGI P2K034 | + | + | ‒ | ‒ | + | + | ‒ | ‒ | ‒ |
| EGI P2K035 | + | ‒ | + | +++ | + | + | + | ‒ | ‒ |
| EGI P2K039 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2K040 | + | ‒ | + | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K041 | ‒ | ‒ | + | ‒ | ‒ | + | + | + | ‒ |
| EGI P2K043 | ‒ | + | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P2K047 | + | + | ++ | ++ | + | + | + | ‒ | ‒ |
| EGI P2K050 | ‒ | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2K052 | ‒ | + | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P2K053 | + | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K054 | + | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2K055 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2K059 | + | + | ‒ | +++ | ‒ | + | + | ‒ | ‒ |
| EGI P2K060 | ‒ | + | + | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K061 | ‒ | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K064 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P2K068 | ‒ | + | + | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K071 | + | ‒ | ‒ | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P2K074 | ‒ | + | ‒ | + | ‒ | + | + | + | ‒ |
| EGI P2K075 | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P2K076 | + | + | ‒ | +++ | ‒ | + | + | + | ‒ |
| EGI P2S006 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2S007 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2S008 | + | + | + | +++ | + | + | + | + | ‒ |
| EGI P2S009 | + | + | ‒ | ++ | + | + | + | + | ‒ |
| EGI P2S010 | ‒ | ‒ | + | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2S013 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2S019 | ‒ | + | + | ‒ | + | + | ‒ | ‒ | ‒ |
| EGI P2S020 | ‒ | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P2S022 | ‒ | + | ‒ | +++ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P2S024 | + | + | ‒ | +++ | + | + | + | + | + |
| EGI P2S027 | + | + | + | ++ | ‒ | + | + | + | ‒ |
| EGI P2S034 | ‒ | + | + | +++ | + | + | + | + | ‒ |
| EGI P2S036 | ‒ | ++ | ‒ | ‒ | + | + | ‒ | ‒ | ‒ |
| EGI P2S040 | ‒ | ‒ | ++ | ‒ | ‒ | + | + | + | + |
| EGI P2S043 | ‒ | + | + | ‒ | + | + | + | + | ‒ |
| EGI P2S045 | ‒ | + | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P2S051 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P2S052 | + | + | + | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P3B002 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3B004 | + | + | + | + | + | + | + | + | ‒ |
| EGI P3B010 | ‒ | + | +++ | ‒ | + | + | + | + | ‒ |
| EGI P3B011 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3B014 | ‒ | ‒ | +++ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3B023 | ‒ | ‒ | ‒ | +++ | ‒ | + | + | ‒ | ‒ |
| EGI P3B024 | ‒ | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P3B025 | + | + | + | ++ | + | + | + | + | ‒ |
| EGI P3B026 | + | + | ++ | +++ | + | + | ‒ | ‒ | ‒ |
| EGI P3B033 | + | + | ‒ | ++ | + | + | + | ‒ | ‒ |
| EGI P3B035 | + | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P3B037 | ‒ | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P3B039 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3B040 | + | ‒ | ‒ | ‒ | + | + | + | + | + |
| EGI P3B044 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P3B047 | + | + | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P3B048 | + | ‒ | + | ++ | + | + | + | + | ‒ |
| EGI P3B055 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P3B056 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P3B057 | ‒ | + | ‒ | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P3B058 | + | ‒ | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P3K001 | + | ++ | + | + | + | + | + | + | ‒ |
| EGI P3K005 | ‒ | ‒ | ‒ | +++ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P3K007 | + | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P3K008 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P3K009 | ‒ | ‒ | ‒ | +++ | + | + | + | ‒ | ‒ |
| EGI P3K010 | + | ‒ | +++ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P3K011 | ‒ | ‒ | + | ‒ | ‒ | + | + | + | ‒ |
| EGI P3K014 | ‒ | ‒ | ‒ | + | ‒ | + | + | ‒ | ‒ |
| EGI P3K015 | + | ‒ | ‒ | ‒ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P3K016 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3K021 | + | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3K023 | + | ++ | + | ‒ | + | + | + | + | ‒ |
| EGI P3K026 | + | + | ‒ | + | + | + | + | ‒ | ‒ |
| EGI P3K027 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3K030 | + | + | + | + | + | + | + | ‒ | ‒ |
| EGI P3K034 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P3K035 | + | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3K036 | ‒ | + | ++ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3K041 | + | + | ‒ | +++ | ‒ | + | ‒ | ‒ | ‒ |
| EGI P3K045 | + | ‒ | ‒ | ‒ | ‒ | + | + | + | + |
| EGI P3K046 | ‒ | + | ‒ | ‒ | + | + | + | + | + |
| EGI P3K047 | ‒ | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P3K051 | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P3K056 | + | ‒ | + | + | + | + | + | + | ‒ |
| EGI P3K057 | ‒ | ‒ | ‒ | ‒ | + | + | + | + | ‒ |
| EGI P3K058 | ‒ | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P3K059 | ‒ | ‒ | + | ‒ | ‒ | + | + | + | ‒ |
| EGI P3S001 | + | + | + | + | + | + | + | + | ‒ |
| EGI P3S003 | + | ‒ | + | ‒ | + | + | + | + | ‒ |
| EGI P3S005 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P3S009 | + | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P3S010 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3S014 | + | ‒ | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3S015 | + | + | + | + | + | + | + | + | ‒ |
| EGI P3S024 | ‒ | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P3S026 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
| EGI P3S031 | ‒ | + | ‒ | ‒ | + | + | + | ‒ | ‒ |
| EGI P3S037 | ‒ | + | + | ‒ | ‒ | + | + | + | ‒ |
| EGI P3S039 | ‒ | ‒ | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P3S042 | ‒ | + | ‒ | ‒ | ‒ | + | + | ‒ | ‒ |
| EGI P3S046 | ‒ | + | ‒ | ‒ | ‒ | + | + | + | ‒ |
Note: ‒, negative (E=1); +, weak (1<E≤2); ++, moderate (2<E≤3); +++, strong (E>3). The ability to nitrogen fixation and salt-tolerant was recorded as ''+'' if strain grows on the test media. E is the ability to solubilize phosphate and produce siderophores, protease, and cellulose. |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
Razzaghi Komaresofla B,
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}