
平茬对天山北坡浅山带灌木群落生长及碳收支的影响
|
刘鑫(1998-),男,硕士研究生,主要从事森林生态系统碳交换研究. E-mail: lx779228239@163.com |
收稿日期: 2025-01-02
修回日期: 2025-03-26
网络出版日期: 2026-03-12
基金资助
新疆维吾尔自治区自然科学基金项目(2023D01A90)
新疆维吾尔自治区自然科学基金项目(2022D01B174)
Effects of levelling stubble on the growth of shrub communities and carbon sequestration in the shallow mountain belt of the northern slope of Tianshan Mountains
Received date: 2025-01-02
Revised date: 2025-03-26
Online published: 2026-03-12
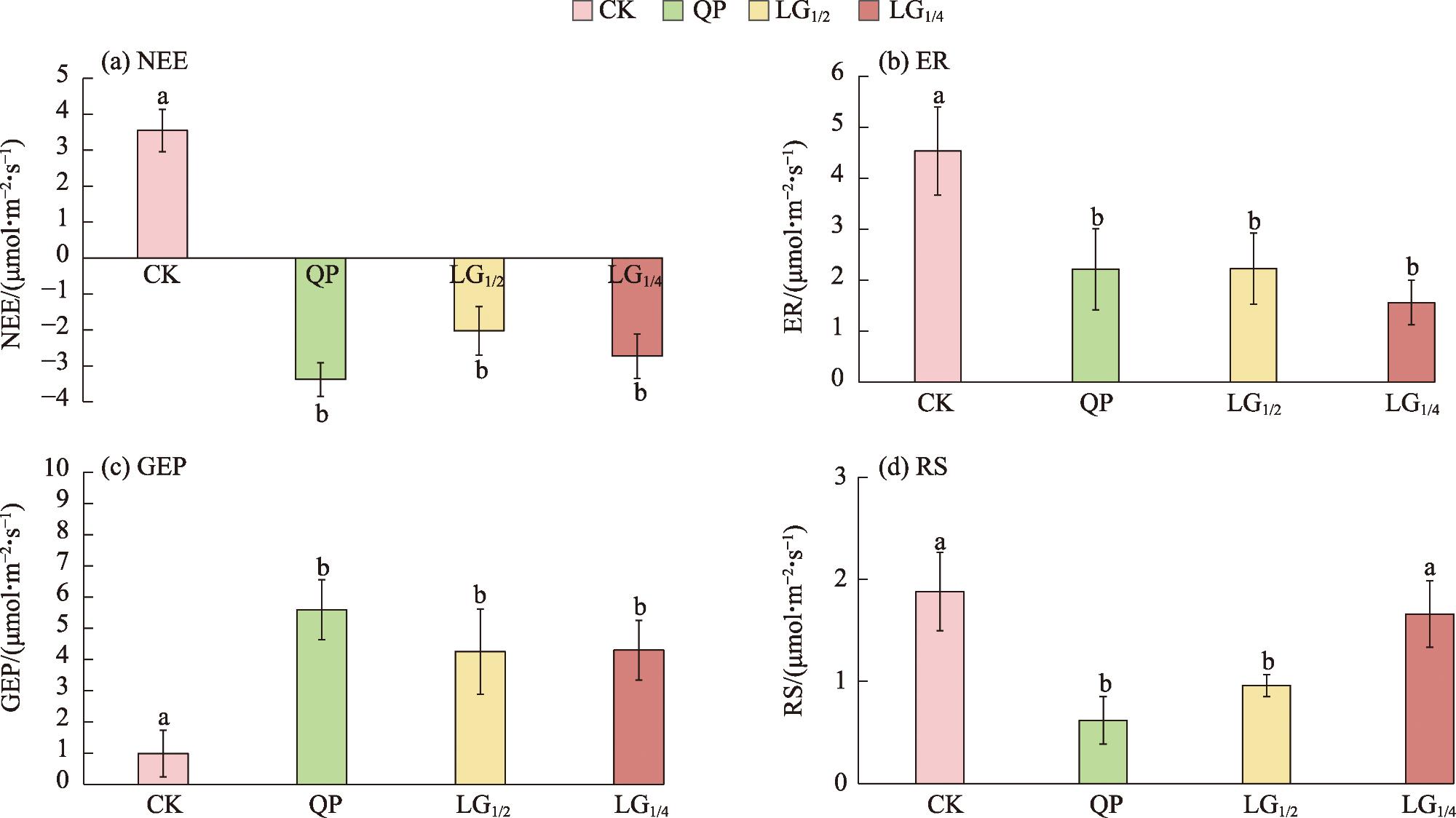
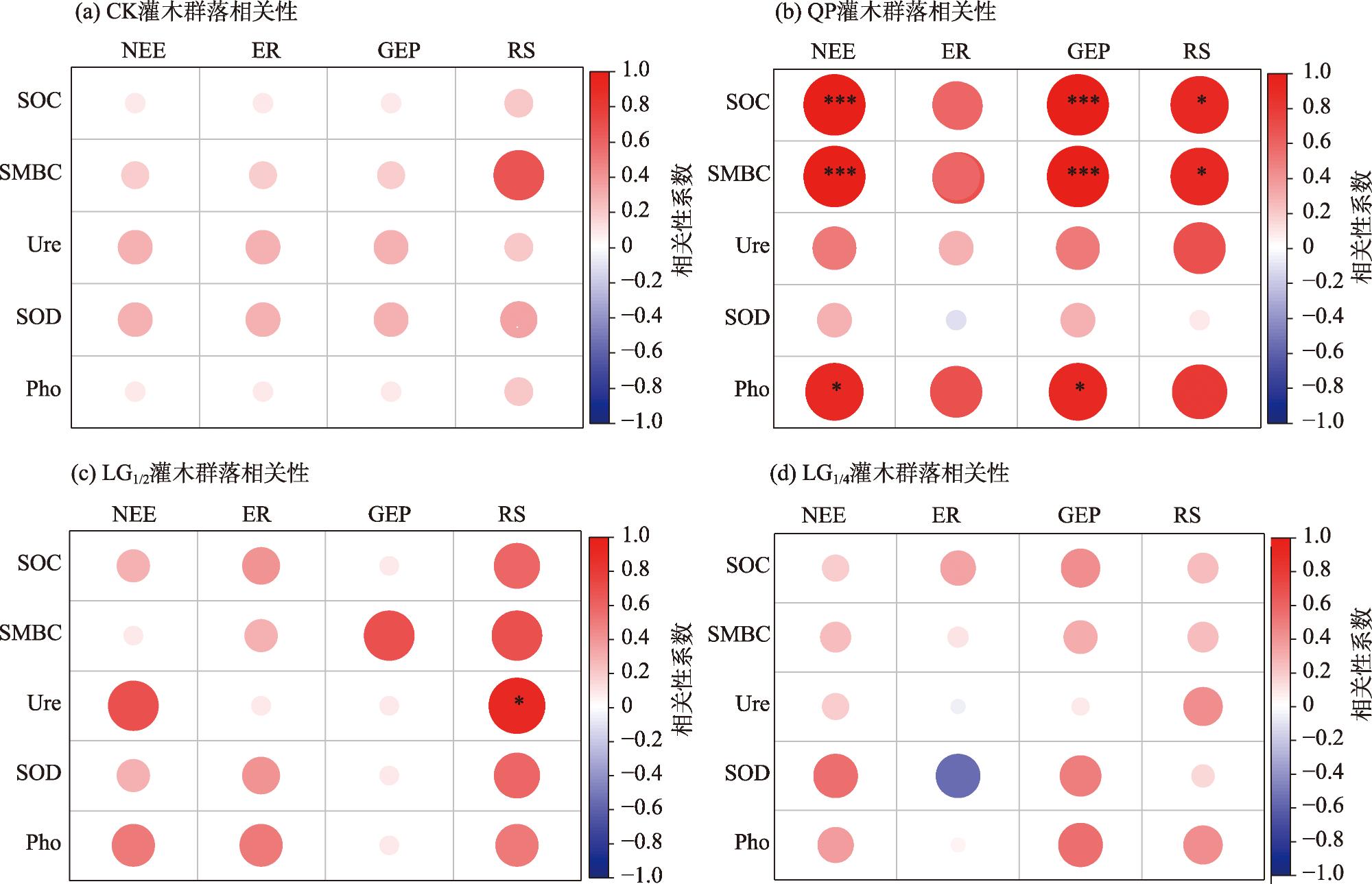
本研究旨在探讨平茬措施对天山北坡浅山带灌木群落生长及碳收支能力的影响。在黑果小檗(Berberis atrocarpa)、宽刺蔷薇(Rosa platyacantha)和金丝桃叶绣线菊(Spiraea hypericifolia)灌木群落内,分别设置4种不同平茬高度处理:全部平茬(QP)、留茬高度为株体原高度的1/2(LG1/2)、留茬高度为株体原高度1/4(LG1/4)、未平茬对照(CK),监测灌木群落生长指标(地径、冠幅、枯枝比、新枝长)、碳收支及土壤微生物和酶活性变化。研究结果表明:(1) 灌木群落的生长指标对QP处理响应最为明显,新枝高度增加109.2%并且枯枝比降幅达到88.1%,QP处理的差异性显著增加。(2) QP、LG1/2、LG1/4平茬处理分别提高了灌木群落的净生态系统CO2交换(NEE)195.3%、157.6%、177.4%,分别降低生态系统呼吸(ER)51.2%、51.7%、66.5%,降低了土壤呼吸(RS)67.4%、49.6%、12.2%,3种平茬处理的灌木群落碳交换均与对照呈现出显著性差异。(3) 平茬处理明显提高灌木群落土壤碳含量和酶活性,土壤碳交换与土壤生物因子(微生物量碳、土壤有机碳、土壤酶活性)显著正相关。综上所述,合理的平茬措施能有效提升天山北坡浅山带灌木群落的生长及固碳能力,全部平茬在该区域内对促进灌木群落生长和提高碳汇功能最为有效,对减缓区域大气中CO2的增长具有积极作用。
刘鑫 , 张毓涛 , 师庆东 , 李吉玫 , 孙雪娇 . 平茬对天山北坡浅山带灌木群落生长及碳收支的影响[J]. 干旱区研究, 2025 , 42(5) : 895 -906 . DOI: 10.13866/j.azr.2025.05.12
This study aims to investigate the effects of stumping on the growth and carbon sequestration capacity of shrub communities in the low mountain belt on the northern slope of the Tianshan Mountains. Within the shrub communities of Berberis atrocarpa, Rosa platyacantha and Spiraea hypericifolia, four different stumping height treatments were set up: complete stumping (QP), leaving 1/2 of the original plant height (LG1/2), leaving 1/4 of the original plant height (LG1/4), and an un-coppiced control (CK). The growth indices of the shrub communities (ground diameter, crown width, dead branch ratio, and new branch length), carbon sequestration, and changes in soil microbial and enzyme activities were monitored. The results showed that: (1) The growth indices of the shrub communities were most responsive to the QP treatment, with new branch height increasing by 109.2% and the dead branch ratio decreasing by 88.1%, resulting in significant differences in the QP treatment. (2) The QP, LG1/2, and LG1/4 treatments increased the net ecosystem CO2 exchange (NEE) of the shrub communities by 195.3%, 157.6%, and 177.4%, respectively, and decreased ecosystem respiration (ER) by 51.2%, 51.7%, and 66.5%, respectively, and soil respiration (RS) by 67.4%, 49.6%, and 12.2%, respectively. The carbon exchange of the shrub communities under the three stumping treatments showed significant differences compared with the control. (3) stumping significantly increased soil carbon content and enzyme activity in the shrub communities, and soil carbon exchange was significantly positively correlated with soil biotic factors (microbial biomass carbon, soil organic carbon, and soil enzyme activity). In conclusion, rational stumping measures can effectively enhance the growth and carbon sequestration capacity of shrub communities in the low mountain belt on the northern slope of the Tianshan Mountains. Complete stumping (QP) is the most effective in promoting shrub community growth and enhancing carbon sink function in this area, and it has a positive effect on mitigating the increase of CO2 in the regional atmosphere.
Key words: carbon balance; flat stubble; shrub communities; net ecosystem; CO2 exchange
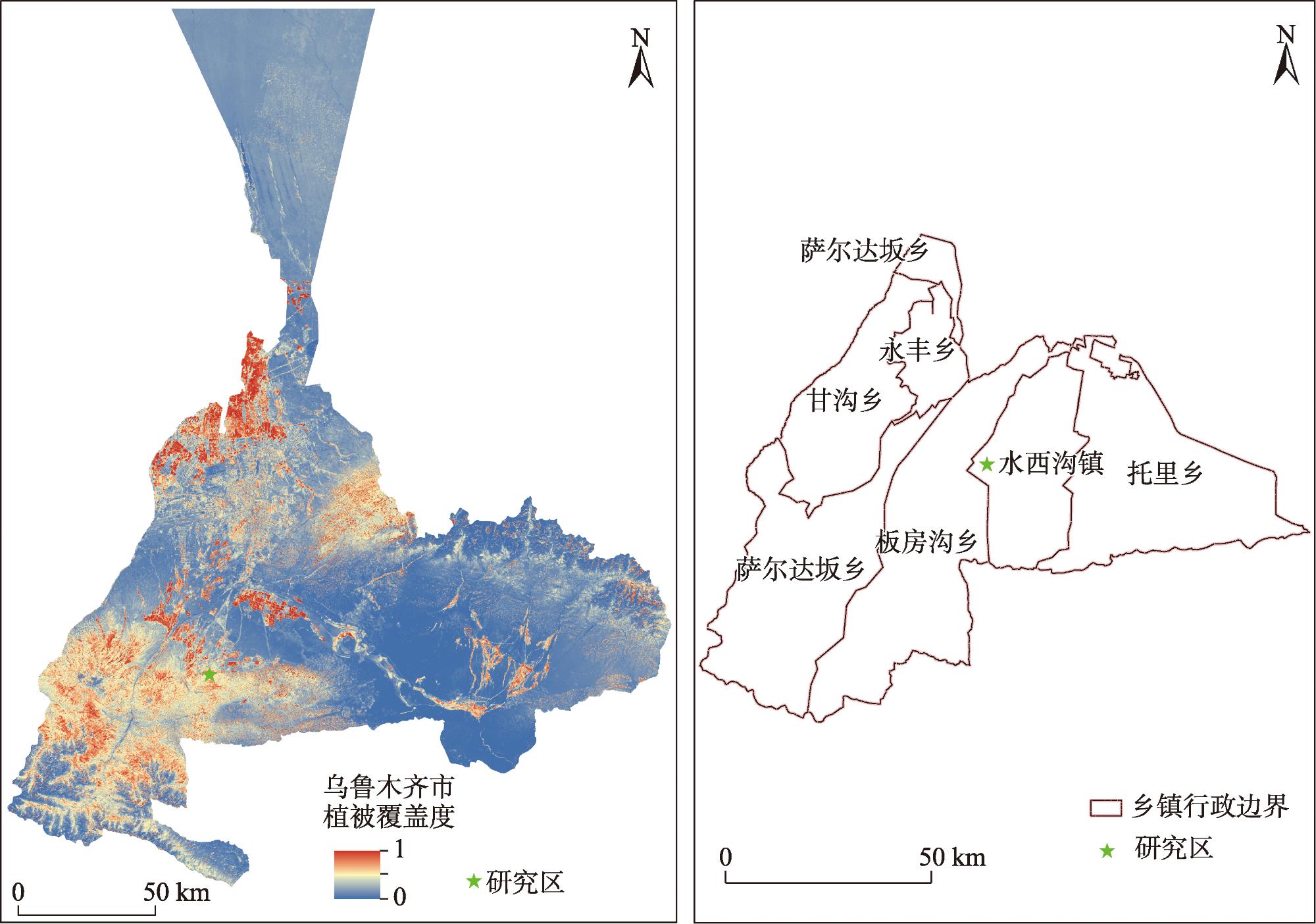
表1 样地基本信息Tab. 1 Basic information on sample plots |
| 平茬处理 | 群落组成 | 样地数量/个 | 灌木平均高度/m | 灌木平均冠幅/m2 | 平均盖度/% | 各样地平均株数/株 |
|---|---|---|---|---|---|---|
| CK | Sh 7株;Rp 8株;Ba 5株 | 4 | 0.42±0.04 | 0.71×0.64 | 34.07±0.02 | 19 |
| QP | Sh 8株;Rp 8株;Ba 4株 | 6 | 0.55±0.06 | 0.59×0.54 | 34.02±0.08 | 26 |
| LG1/2 | Sh 4株;Rp 9株;Ba 7株 | 4 | 0.69±0.09 | 0.71×0.68 | 41.13±0.10 | 26 |
| LG1/4 | Sh 5株;Rp 8株;Ba 7株 | 4 | 0.85±0.08 | 0.60×0.54 | 40.05±0.11 | 30 |
注:CK为对照组;QP为全部平茬(留茬高度3~5 cm);LG1/2为平茬高度为株体原高度的1/2;LG1/4为平茬高度为株体原高度1/4;Sh为金丝桃叶绣线菊;Rp为宽刺蔷薇;Ba为黑果小檗。下同。 |
表2 同化箱内样株基本情况Tab. 2 Basic information on the sample plants in the assimilation chamber |
| 处理 | 同化箱内树种数量及组成 | 平均株高/m | 平均冠幅/m2 | 平均盖度/% |
|---|---|---|---|---|
| CK | Sh 2株;Rp 1株;Ba 1株 | 0.38±0.14 | 0.35±0.07 | 43.10±0.07 |
| QP | Sh 1株;Rp 1株;Ba 1株 | 0.42±0.06 | 0.32±0.06 | 45.06±0.06 |
| LG1/2 | Sh 1株;Rp 1株;Ba 1株 | 0.50±0.17 | 0.37±0.15 | 47.06±0.05 |
| LG1/4 | Sh 1株;Rp 1株;Ba 1株 | 0.65±0.26 | 0.32±0.09 | 50.03±0.11 |
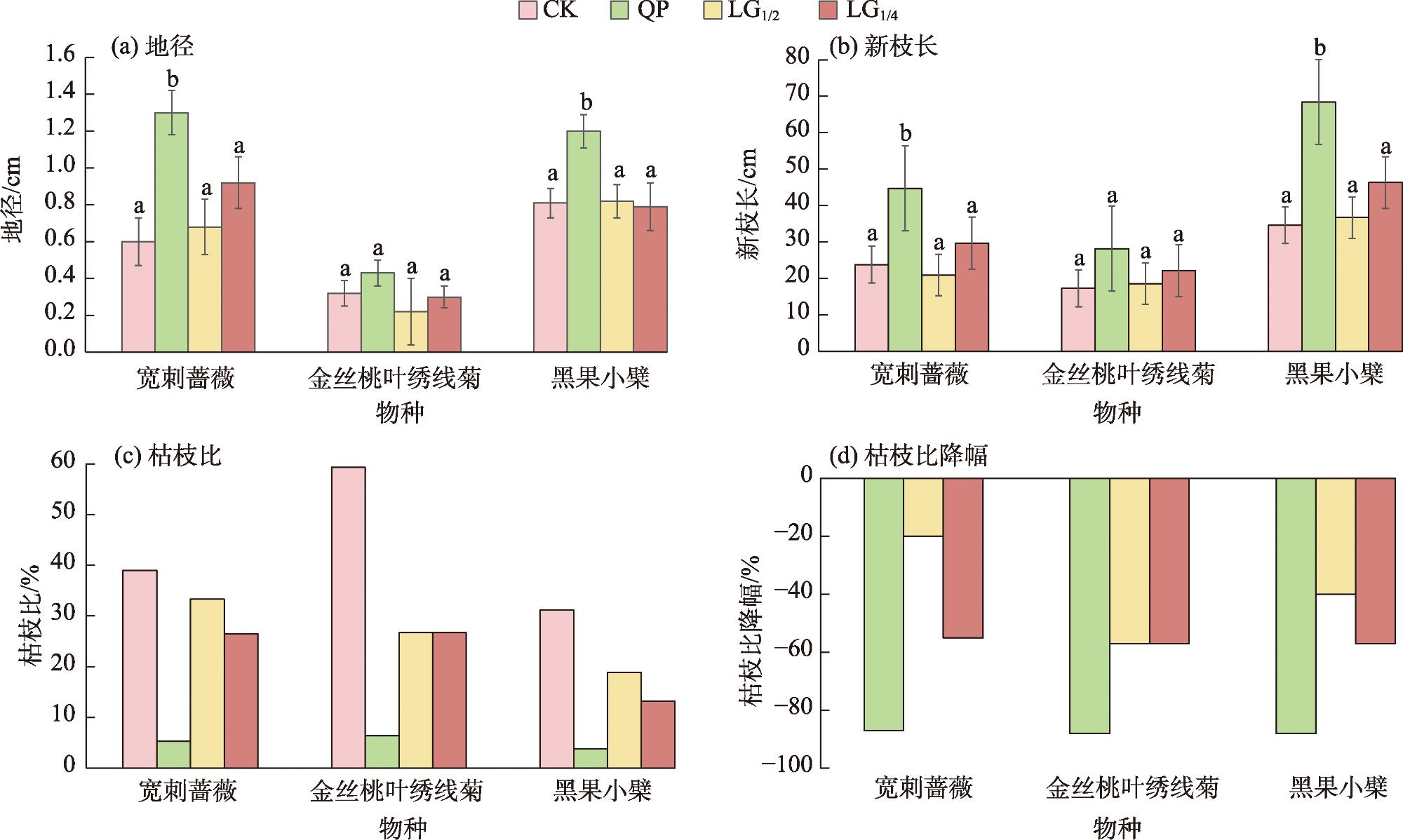
表3 不同平茬高度3种灌木地径变化率Tab. 3 Rates of change in ground diameter of three shrubs with different levelling heights |
| 物种 | QP | LG1/2 | LG1/4 |
|---|---|---|---|
| 宽刺蔷薇 | 116.7 | 5.0 | 50.0 |
| 金丝桃叶绣线菊 | 33.3 | -33.3 | -6.7 |
| 黑果小檗 | 50.0 | 2.5 | -2.5 |
注:以上均为与CK相比的变化率(%)。 |
表4 不同平茬高度3种灌木新枝长与变化率Tab. 4 New branch length and rates of change of three shrubs with different levelling heights % |
| 指标 | 处理 | 物种 | ||
|---|---|---|---|---|
| 宽刺蔷薇 | 金丝桃叶绣线菊 | 黑果小檗 | ||
| 新枝长/cm | CK | 23.0 | 18.0 | 35.0 |
| QP | 44.0 | 28.0 | 73.0 | |
| LG1/2 | 21.0 | 20.0 | 38.0 | |
| LG1/4 | 29.0 | 22.0 | 48.0 | |
| 平茬处理与CK的新枝长度变化率/% | QP与CK | 91.2 | 56.5 | 109.2 |
| LG1/2与CK | -9.3 | 11.4 | 9.1 | |
| LG1/4与CK | 26.4 | 22.1 | 37.1 | |
表5 不同平茬高度3种灌木单株萌蘖枝条数与变化率Tab. 5 Number and rate of change of sprouting tillers of three shrubs at different levelling heights % |
| 指标 | 处理 | 物种 | ||
|---|---|---|---|---|
| 宽刺蔷薇 | 金丝桃叶绣线菊 | 黑果小檗 | ||
| 单株萌蘖枝条 数/个 | CK | 3 | 12 | 5 |
| QP | 10 | 28 | 21 | |
| LG1/2 | 5 | 16 | 13 | |
| LG1/4 | 8 | 18 | 16 | |
| 平茬处理与CK的单株萌蘖枝条数变化率/% | QP与CK | 233.1 | 133.2 | 323.0 |
| LG1/2与CK | 67.5 | 33.1 | 164.0 | |
| LG1/4与CK | 167.3 | 50.3 | 224.0 | |
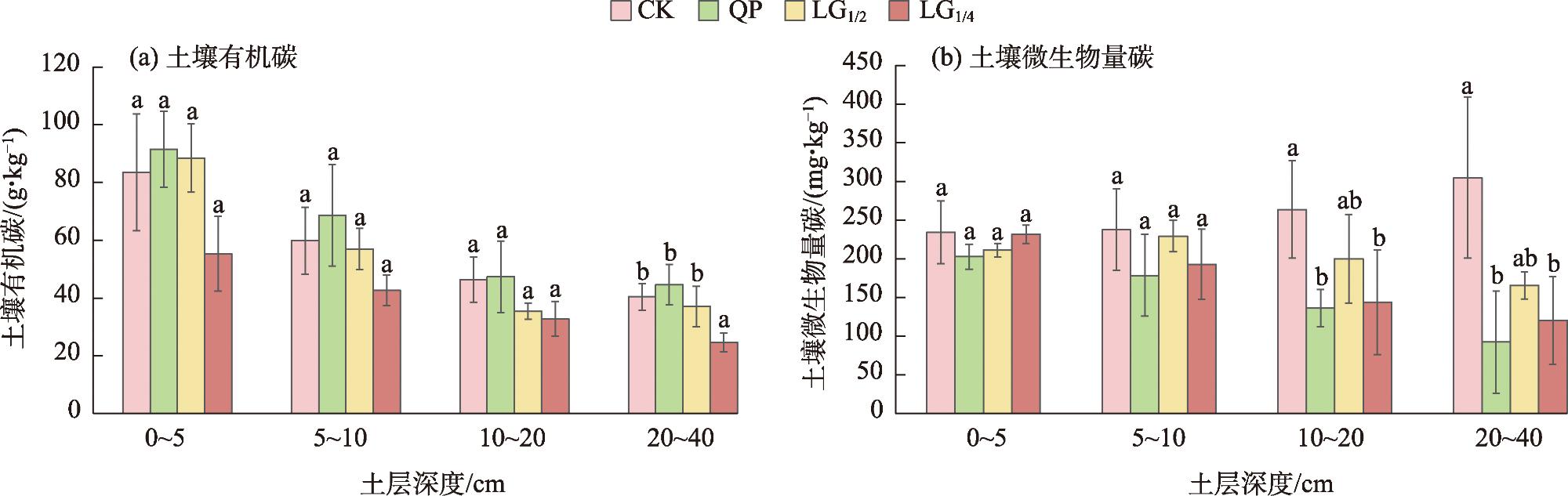
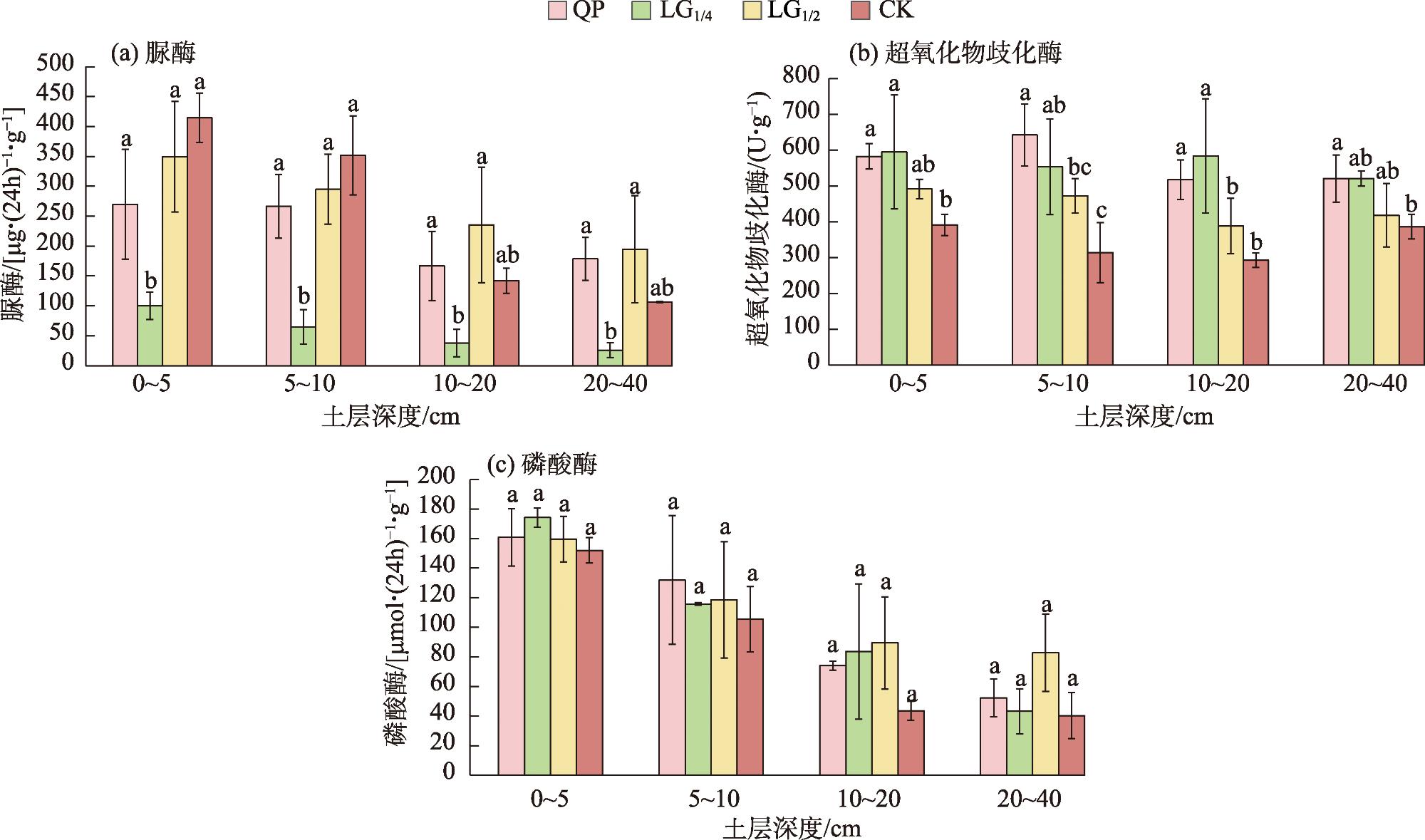
表6 灌木群落土壤碳及酶活性垂直变化Tab. 6 Vertical changes in soil carbon and enzyme activity in shrub communities |
| 平茬处理 | 土层深度 /cm | 土壤有机碳 /(g·kg-1) | 土壤微生物量碳 /(mg·kg-1) | 脲酶 /[μg·(24h)-1·g-1] | 超氧化物歧化酶 /(U·g-1) | 磷酸酶 /[μmol·(24h)-1·g-1] |
|---|---|---|---|---|---|---|
| CK | 0~5 | 55.27±12.94 | 231.53±11.82 | 414.45±41.41 | 391.35±29.30 | 151.98±8.57 |
| 5~10 | 42.60±5.32 | 192.63±45.39 | 351.52±66.13 | 314.03±83.91 | 105.48±22.06 | |
| 10~20 | 32.78±6.07 | 144.03±67.68 | 142.20±21.01 | 292.98±20.20 | 43.57±6.38 | |
| 20~40 | 24.61±3.23 | 120.35±56.55 | 106.37±1.24 | 386.52±34.32 | 40.31±11.50 | |
| QP | 0~5 | 83.48±20.20 | 234.19±40.44 | 269.80±71.74 | 582.74±35.93 | 160.85±19.42 |
| 5~10 | 59.81±11.66 | 237.52±52.75 | 266.60±53.60 | 642.53±86.23 | 131.95±43.55 | |
| 10~20 | 46.39±7.85 | 263.53±62.87 | 166.67±57.96 | 517.83±55.37 | 84.08±3.10 | |
| 20~40 | 40.43±4.60 | 304.77±74.14 | 178.62±36.06 | 521.14±65.67 | 52.31±12.91 | |
| LG1/4 | 0~5 5~10 10~20 20~40 | 91.49±13.22 68.66±17.61 47.34±12.37 44.60±7.05 | 202.62±16.26 178.55±52.82 136.12±20.41 92.30±25.39 | 99.81±22.87 64.85±19.05 37.76±10.87 25.96±7.36 | 595.02±158.70 553.41±133.68 583.83±159.68 520.49±21.04 | 174.18±6.62 115.82±0.89 83.68±25.72 43.11±12.22 |
| LG1/2 | 0~5 5~10 10~20 20~40 | 88.43±11.84 56.97±7.17 35.41±2.83 37.03±7.02 | 211.14±8.71 229.16±20.41 200.02±57.39 165.33±17.29 | 349.34±52.66 294.86±58.32 235.72±56.56 194.64±59.61 | 491.81±26.64 472.25±48.15 388.04±77.31 418.56±88.58 | 159.58±15.38 118.56±35.37 89.44±21.12 82.89±16.21 |
图4 不同平茬高度下灌木群落土壤有机碳、土壤微生物量碳含量变化Fig. 4 Changes in soil organic carbon and soil microbid biomass carbon of shrub communities under different levelling height |
| [1] |
张颖, 李晓格, 温亚利. 碳达峰碳中和背景下中国森林碳汇潜力分析研究[J]. 北京林业大学学报, 2022, 44(1): 38-47.
[
|
| [2] |
|
| [3] |
付玉杰, 田地, 侯正阳, 等. 全球森林碳汇功能评估研究进展[J]. 北京林业大学学报, 2022, 44(10): 1-10.
[
|
| [4] |
朴世龙, 岳超, 丁金枝, 等. 试论陆地生态系统碳汇在“碳中和”目标中的作用[J]. 中国科学: 地球科学, 2022, 52(7): 1419-1426.
[
|
| [5] |
王兴昌, 王传宽, 于贵瑞. 基于全球涡度相关的森林碳交换的时空格局[J]. 中国科学(D辑:地球科学), 2008, 38(9): 1092-1102.
[
|
| [6] |
周丽艳, 贾丙瑞, 周广胜, 等. 中国北方针叶林生长季碳交换及其调控机制[J]. 应用生态学报, 2010, 21(10): 2449-2456.
[
|
| [7] |
陈骥. 模拟增温和围栏封育对青海湖北岸高寒草甸化草原生态系统碳交换影响[D]. 西安: 中国科学院研究生院(地球环境研究所), 2015.
[
|
| [8] |
方显瑞. 杨树人工林生态系统碳交换及其环境响应[D]. 北京: 北京林业大学, 2011.
[
|
| [9] |
李金山, 马旭君, 吴晶, 等. 不同强度平茬对柠条生长及养分归还的影响[J]. 西部林业科学, 2023, 52(1): 57-63.
[
|
| [10] |
李宇, 徐新文, 许波, 等. 塔里木沙漠公路防护林乔木状沙拐枣平茬复壮技术的研究[J]. 干旱区资源与环境, 2014, 28(2): 103-108.
[
|
| [11] |
裴志永, 段广东, 郝少荣, 等. 基于防风效益的毛乌素沙地沙柳灌丛平茬方法优选[J]. 农业工程学报, 2019, 35(4): 153-161.
[
|
| [12] |
尼珍, 赵垦田, 杨小林, 等. 平茬高度对砂生槐萌蘖的影响[J]. 林业调查规划, 2014, 39(3): 114-116.
[
|
| [13] |
|
| [14] |
|
| [15] |
张泽宁, 李芳, 郭彩云, 等. 中国沙棘伐桩萌枝能力对平茬高度的响应[J]. 西南林业大学学报(自然科学), 2020, 40(6): 34-39.
[
|
| [16] |
聂恺宏, 吉生丽, 邹旭, 等. 中国沙棘平茬萌蘖动态及其对种群结构的影响[J]. 云南大学学报(自然科学版), 2018, 40(4): 804-813.
[
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
田登娟, 白双成, 聂恺宏, 等. 平茬高度对中国沙棘萌枝能力及非结构性碳水化合物积累与分配的影响[J]. 西北植物学报, 2021, 41(4): 627-634.
[
|
| [21] |
于文涛. 平茬措施对柠条生理特征及土壤理化性质的影响[D]. 杨凌: 西北农林科技大学, 2016.
[
|
| [22] |
董雪, 郝玉光, 辛智鸣, 等. 科尔沁沙地4种典型灌木灌丛下土壤碳、氮、磷化学计量特征[J]. 西北植物学报, 2019, 39(1): 164-172.
[
|
| [23] |
李元, 时伟宇, 闫美杰, 等. 土壤呼吸影响因素概述及展望[J]. 水土保持研究, 2013, 20(5): 311-316.
[
|
| [24] |
|
| [25] |
谢宗强, 唐志尧. 中国灌丛生态系统碳储量的研究[J]. 植物生态学报, 2017, 41(1): 1-4.
[
|
| [26] |
陈晓阳. 中国灌木林资源[M]. 北京: 中国林业出版社, 2021.
[
|
| [27] |
肖媛媛, 冯薇, 乔艳桂, 等. 固沙灌木林地土壤微生物群落特征对土壤多功能性的影响[J]. 生物多样性, 2023, 31(4): 128-141.
[
|
| [28] |
李文, 李小龙, 刘玉祯, 等. 不同强度放牧对东祁连山高寒灌丛土壤理化特征的影响[J]. 草原与草坪, 2020, 40(4): 8-15.
[
|
| [29] |
刘启嵘. 平茬对沙柳生长及碳汇能力的影响[D]. 呼和浩特: 内蒙古农业大学, 2022.
[
|
| [30] |
骆土寿, 陈步峰, 李意德, 等. 海南岛尖峰岭热带山地雨林土壤和凋落物呼吸研究[J]. 生态学报, 2001, 21(12): 2013-2017.
[
|
| [31] |
李耀林, 郭忠升. 平茬对半干旱黄土丘陵区柠条林地土壤水分的影响[J]. 生态学报, 2011, 31(10): 2727-2736.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}