
毛乌素沙地不同立地条件沙柳生物量异速生长规律
|
王岳(1989-),男,博士,副教授,主要从事沙区资源高效利用等方面研究. E-mail: wyue@imufe.edu.cn |
收稿日期: 2024-06-23
修回日期: 2024-12-10
网络出版日期: 2025-08-13
基金资助
内蒙古自治区科技计划项目(2023YFHH0068)
国家自然科学基金项目(31700439)
国家自然科学基金项目(U23A2014)
内蒙古自治区“五大任务”研究专项(NCXWD2472)
Biomass allocation in Salix psammophila and the response to different site conditions in the Mu Us Sandy Land
Received date: 2024-06-23
Revised date: 2024-12-10
Online published: 2025-08-13
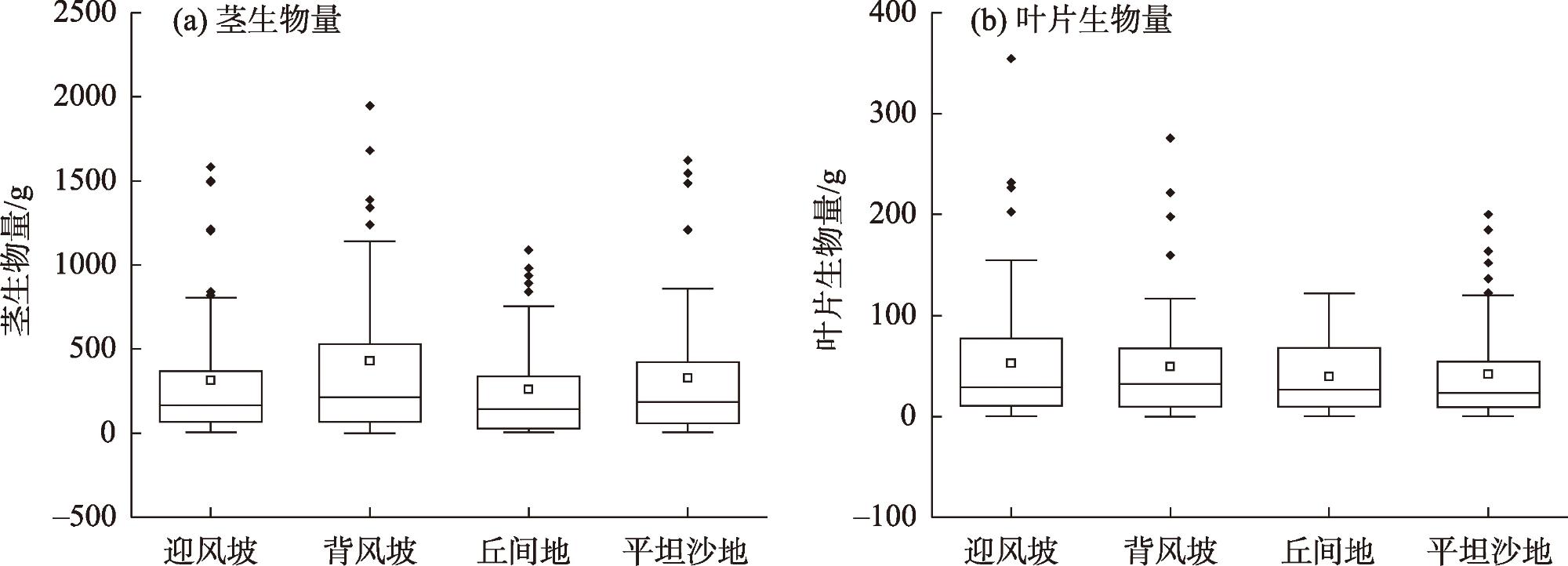
为从生物量积累和分配的角度探究沙柳(Salix psammophila)对不同沙地环境条件的适应策略,揭示影响沙柳生物量分配的主要环境因子,为沙柳种群稳定性建设和可持续经营提供参考,选取沙丘迎风坡、背风坡、丘间地和平坦沙地4种立地类型的沙柳林为研究对象,测定各样地内沙柳代表株的丛生枝基径、长度和茎叶生物量,用标准主轴回归建立茎、叶生物量与基径、枝长的异速生长方程,以探明沙柳丛生枝尺度上茎叶生物量积累分配与个体尺寸的依赖关系;进一步通过主轴漂移的方法,检验茎叶生物量异速生长方程在不同立地类型条件下的差异,进而探讨环境条件对沙柳生物量积累和分配的影响。结果表明:(1)随丛生枝个体增大,沙柳茎生物量积累速率高于叶生物量积累速率,这一规律在各个立地类型间表现一致;(2)沙柳丛生枝茎、叶生物量分配在不同立地类型间差异显著,沙丘(迎风坡、背风坡)的叶生物量占丛生枝条生物量的比例显著高于丘间地和平坦沙地(P<0.05);(3)造成沙柳生物量及其分配出现立地类型差异的主要土壤因子是土壤含水率,其中全剖面含水率(0~100 cm)与丛生枝生物量显著相关(P<0.01),深层土壤含水率(60~100 cm)与丛生枝地上生物量分配显著相关。沙柳的生物量积累与分配在不同立地类型间具有显著差异,深层土壤含水率比全剖面土壤含水率对生物量分配的影响更显著,基于本研究结果,未来可以通过适当人工干预以满足沙柳生长的水分要求,实现沙柳人工林种群的稳定发展。
王岳 , 刘中华 , 于明含 , 贺宇 , 郝蕾 , 刘学瑶 , 安骁 . 毛乌素沙地不同立地条件沙柳生物量异速生长规律[J]. 干旱区研究, 2025 , 42(2) : 289 -298 . DOI: 10.13866/j.azr.2025.02.09
This study aimed to clarify the adaptability strategies of Salix psammophila to different environments from the perspective of resource allocation for the stable construction and sustainable management of S. psammophila plantations. S. psammophila plantations in four types of sites (windward slope, leeward slope, interdune land, and flat sand land) were surveyed in the Mu Us Sandy Land in Ningxia. The characteristics of the S. psammophila population, including morphological parameters like basal diameter, branch length, and stem and leaf biomass, were recorded. The effects of topography on the biomass accumulation and distribution between the S. psammophila organs were studied based on the allometric growth model. (1) With growth, S. psammophila tends to reduce the proportion of leaf resources and increase the proportion of stem resources. (2) The site factors significantly impact biomass and its distribution between the S. psammophila organs. The average biomass of branches in the dune (windward and leeward slopes) is significantly higher than that in the interdune land (P<0.05), and the allometric growth index of leaf-stem biomass is significantly higher in the dune than in the interdune and flat sand lands (P<0.05). (3) The main soil factor that affects biomass and its distribution between the S. psammophila organs is soil moisture. Soil moisture of the whole section (0-100 cm) determines the biomass, and soil moisture of the deep layer moisture content (60-100 cm) affects the biomass distribution trade-off between the stem and leaf. The biomass accumulation and allocation of S. psammophila displayed significant differences among different site types, and the effect of deep soil moisture on biomass allocation was more significant than that of full-profile soil moisture. In the future, artificial intervention can be adopted to satisfy the water requirement for S. psammophila growth and realize the stable development of the S. psammophila plantation population.
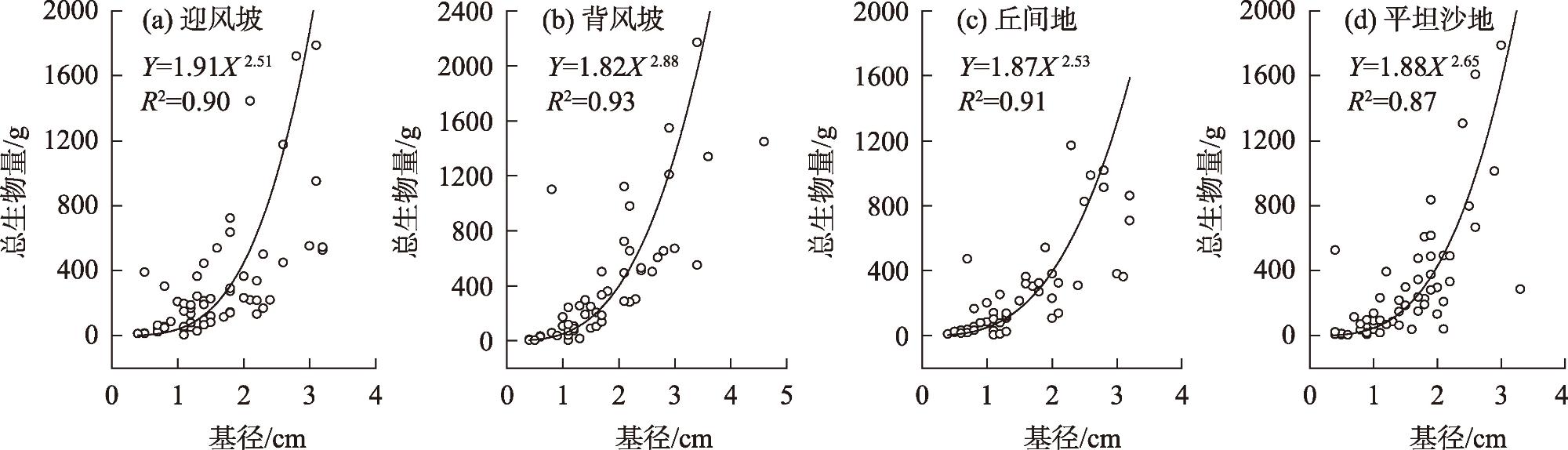
图3 各立地类型沙柳丛生枝地上生物量与基径的关系Fig. 3 Correlation of above-ground biomass and basal diameter for Salix psammophila under different site type |
表1 不同立地类型的沙柳丛生枝生物量-基径之间的异速生长检验以及主轴漂移检验Tab. 1 Allometric relationships, allometry test, and test of the common slope between biomass and basal diameter for Salix psammophila under different site type |
| 参数 | 微环境 | 异速生长检验 | 主轴漂移检验 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| α | logβ | R2 | P | 迎风坡 | 背风坡 | 丘间地 | 平坦沙地 | |||
| Y=B0 X=D | 迎风坡 | 2.51 | 1.91 | 0.90 | <0.01 | 1 | ||||
| 背风坡 | 2.88 | 1.82 | 0.93 | <0.01 | * | 1 | ||||
| 丘间地 | 2.53 | 1.87 | 0.91 | <0.01 | NS | * | 1 | |||
| 平坦沙地 | 2.65 | 1.88 | 0.87 | <0.01 | NS | NS | NS | 1 | ||
| Y=Bs X=D | 迎风坡 | 2.32 | 1.69 | 0.88 | <0.01 | 1 | ||||
| 背风坡 | 2.64 | 1.62 | 0.92 | <0.01 | NS | 1 | ||||
| 丘间地 | 2.58 | 1.61 | 0.93 | <0.01 | NS | NS | 1 | |||
| 平坦沙地 | 2.61 | 1.68 | 0.88 | <0.01 | NS | NS | NS | 1 | ||
| Y=Bl X=D | 迎风坡 | 2.91 | 0.97 | 0.66 | <0.01 | 1 | ||||
| 背风坡 | 3.01 | 0.83 | 0.78 | <0.01 | * | 1 | ||||
| 丘间地 | 2.37 | 1.08 | 0.65 | <0.01 | NS | * | 1 | |||
| 平坦沙地 | 2.56 | 0.94 | 0.72 | <0.01 | NS | NS | NS | 1 | ||
注:*代表在0.05水平显著;**代表在0.01水平显著;NS代表不显著。下同。 |
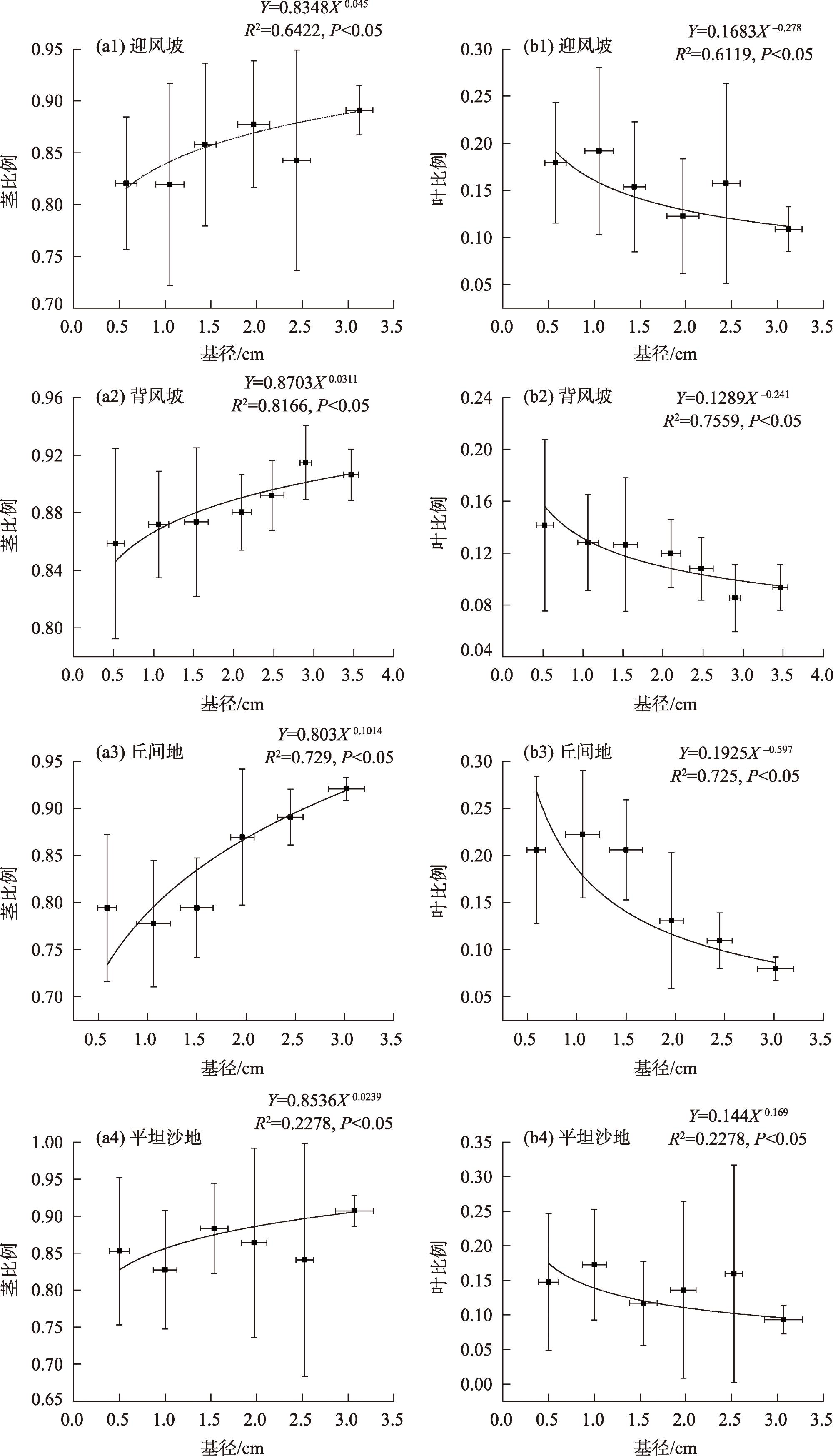
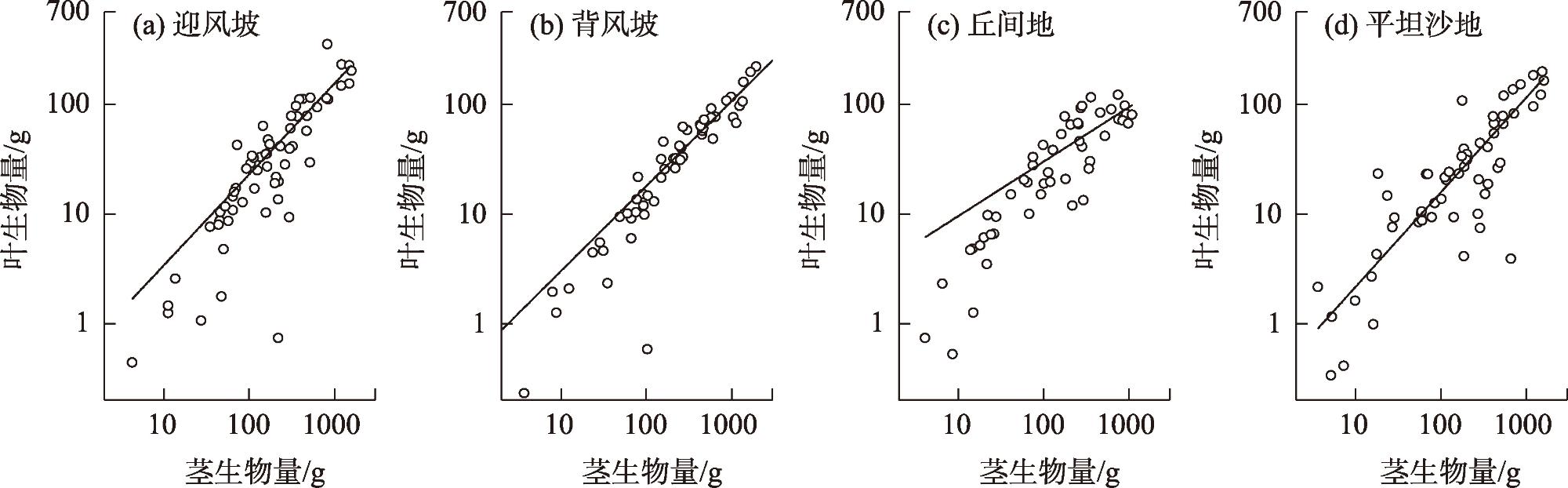
图5 各立地类型下沙柳丛生枝茎-叶生物量的异速生长关系Fig. 5 Allometric relationship between leaf biomass and stem biomass for Salix psammophila branch under different site types |
表2 不同立地类型的沙柳丛生枝茎-叶生物量之间的异速生长检验以及主轴漂移检验Tab. 2 Allometric relationships, allometry test, and test of the common slope between different modules biomass of Salix psammophila under different site types |
| 参数 | 地形 | 异速生长模型拟合结果 | 等速生长检验 | 主轴漂移检验 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| α | logβ | R2 | P | WS | LS | DI | SL | ||||
| Y=Bl X=Bs | 迎风坡 | 0.96 | 0.19 | 0.72 | 0.25 | 1 | |||||
| 背风坡 | 1.05 | 0.09 | 0.88 | 0.13 | NS | 1 | |||||
| 丘间地 | 0.82 | 0.43 | 0.81 | <0.01 | ** | * | 1 | ||||
| 平坦沙地 | 0.81 | 0.34 | 0.71 | <0.01 | ** | * | NS | 1 | |||
注:WS代表迎风坡;LS代表背风坡;DI代表丘间地;SL代表平坦沙地。 |
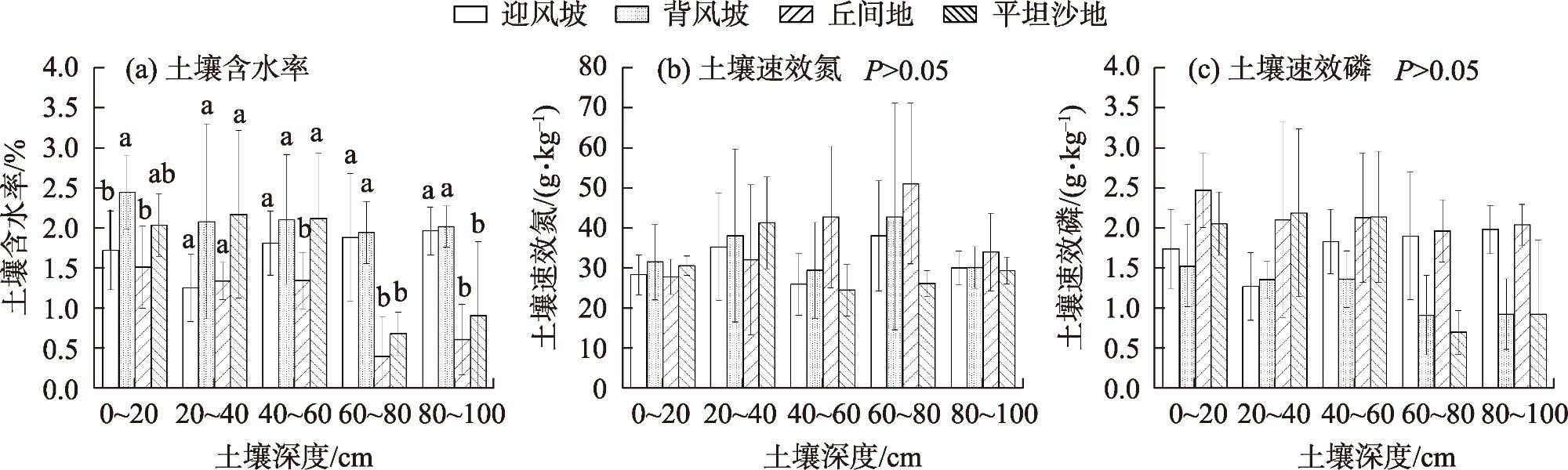
表3 土壤性质与分配特征的相关系数Tab. 3 Correlation coefficients between soil characteristics and allometric index of Bl -Bs |
| 指标 | 土壤深度/cm | 含量 | 叶片-茎干异速生长指数α | 相关性 检验 |
|---|---|---|---|---|
| 土壤含水率/% | 0~20 | 2.48±0.04 | 0.654 | 1.09 |
| 20~40 | 1.84±0.07 | 0.47 | ||
| 40~60 | 1.43±0.09 | 0.89 | ||
| 60~80 | 0.91±0.11 | 0.03* | ||
| 80~100 | 0.42±0.03 | 0.02* | ||
| 土壤速效磷 /(g·kg-1) | 0~20 | 2.365±0.891 | 1.24 | |
| 20~40 | 2.485±0.120 | 1.81 | ||
| 40~60 | 1.762±0.173 | 2.87 | ||
| 60~80 | 0.148±0.059 | 3.46 | ||
| 80~100 | 0.187±0.078 | 1.98 | ||
| 土壤速效氮 /(g·kg-1) | 0~20 | 50.245±0.215 | 2.55 | |
| 20~40 | 41.147±1.037 | 1.86 | ||
| 40~60 | 38.265±1.857 | 3.07 | ||
| 60~80 | 24.344±1.664 | 2.69 | ||
| 80~100 | 27.068±2.078 | 4.28 |
注:*代表P<0.05的显著性。 |
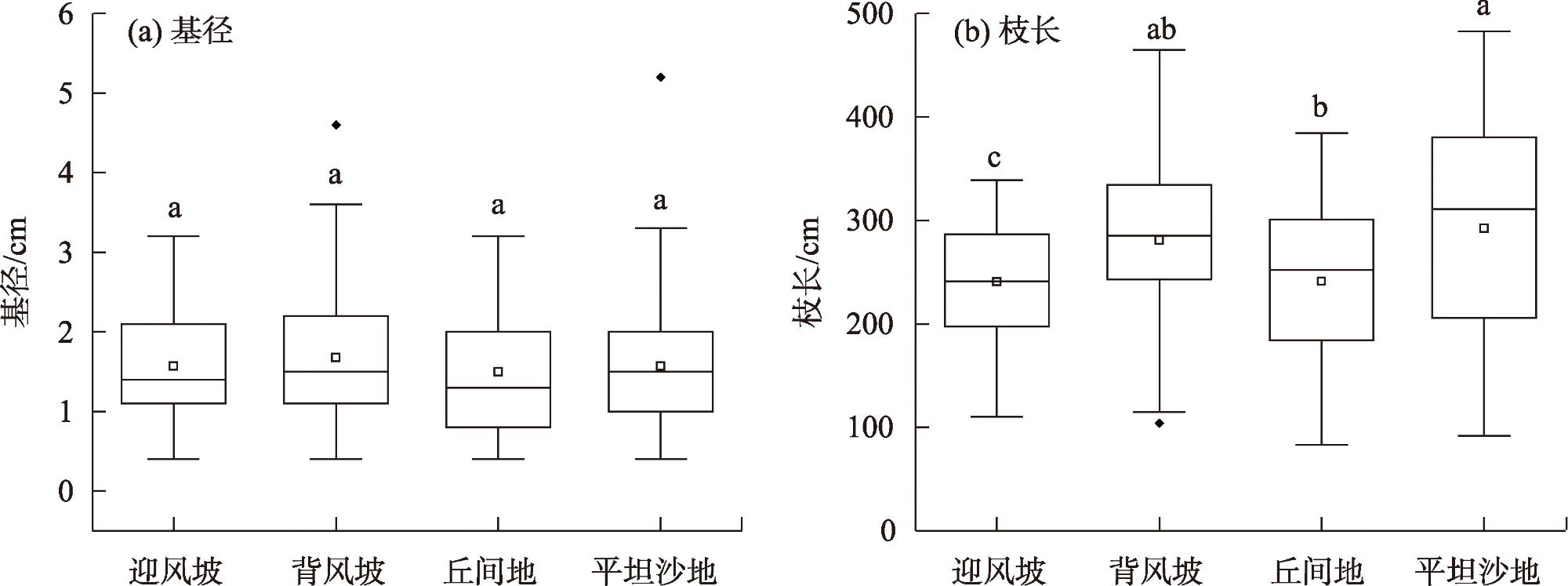
表4 沙柳丛生枝枝长-基径形态参数之间的异速生长检验Tab. 4 Allometric relationships, allometry test between basal diameter and branch lenghth of Salix psammophila |
| 参数 | 异速生长模型拟合结果 | 异速生长检验 | ||||
|---|---|---|---|---|---|---|
| α | logβ | R2 | P | |||
| Y=L | X=D | 0.61 | 2.31 | 0.70 | <0.01 | |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
李雪华, 李晓兰, 蒋德明, 等. 科尔沁沙地70种草本植物个体和构件生物量比较研究[J]. 干旱区研究, 2009, 26(2): 54-59.
[
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
陈国鹏, 赵文智, 何世雄, 等. 沙柳丛生枝生物量最优分配与异速生长[J]. 中国沙漠, 2016, 36(2): 357-363.
[
|
| [9] |
|
| [10] |
|
| [11] |
高凯, 张丽娟, 于永奇, 等. 沙地土壤pH值、养分含量对地形变化的响应[J]. 水土保持通报, 2016, 36(1): 88-92.
[
|
| [12] |
李民青, 周乐, 王喜勇, 等. 7种荒漠木本植物枝干与叶片光合特征及其影响因素[J]. 应用生态学报, 2023, 34(10): 2637-2643.
[
|
| [13] |
|
| [14] |
贾美玉, 李雪华, 吴忠铉, 等. 科尔沁沙地流动沙丘3种常见植物的空间分布格局与异速生长[J]. 应用生态学报, 2015, 26(10): 2953-2960.
[
|
| [15] |
海龙, 王晓江, 张文军, 等. 毛乌素沙地人工沙柳(Salix Psammophila)林平茬复壮技术[J]. 中国沙漠, 2016, 36(1): 131-136.
[
|
| [16] |
李阳, 裴志永, 秦伟, 等. 区域性皆伐抚育作业对沙柳林地土壤养分空间异质性的影响[J]. 科学技术与工程, 2016, 16(18): 161-165.
[
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
张志永, 时忠杰, 张晓, 等. 浑善达克沙地不同地形的土壤物理性质和草本群落分布及其相关性分析[J]. 植物资源与环境学报, 2017, 26(1): 69-76.
[
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
原鹏飞, 丁国栋, 王炜炜, 等. 毛乌素沙地降雨入渗和蒸发特征[J]. 中国水土保持科学, 2008, 6(4): 23-27.
[
|
| [28] |
张军红, 吴波, 杨文斌, 等. 不同演替阶段油蒿群落土壤水分特征分析[J]. 中国沙漠, 2012, 32(6): 1597-1603.
[
|
| [29] |
艾绍水, 李秧秧, 陈佳村, 等. 陕北沙地3种典型灌木根木质部解剖结构及水力特性[J]. 应用生态学报, 2015, 26(11): 3277-3284.
[
|
| [30] |
卜崇峰, 刘国彬, 张文辉. 黄土丘陵沟壑区狼牙刺的生长特征研究[J]. 西北植物学报, 2004, 24(10): 1792-1797.
[
|
| [31] |
陈文思, 朱清科, 刘蕾蕾, 等. 陕北半干旱黄土区沙棘人工林的死亡率及适宜地形因子[J]. 林业科学, 2016, 52(5): 9-16.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}