
基于SSR分子标记的5个黄花补血草居群花型变异及其遗传效应
|
张晶(1996-),女,硕士研究生,主要从事植物繁殖生态学研究. E-mail: 2276834219@qq.com |
收稿日期: 2024-01-09
修回日期: 2024-06-09
网络出版日期: 2025-08-13
基金资助
国家自然科学基金项目(32360308)
自治区教育厅项目(XJEDU2021I006)
Floral morph variation and genetic effect of five Limonium aureum populations based on Simple Sequence Repeat molecular markers
Received date: 2024-01-09
Revised date: 2024-06-09
Online published: 2025-08-13
张晶 , 何爽 , 张爱勤 . 基于SSR分子标记的5个黄花补血草居群花型变异及其遗传效应[J]. 干旱区研究, 2025 , 42(3) : 499 -510 . DOI: 10.13866/j.azr.2025.03.10
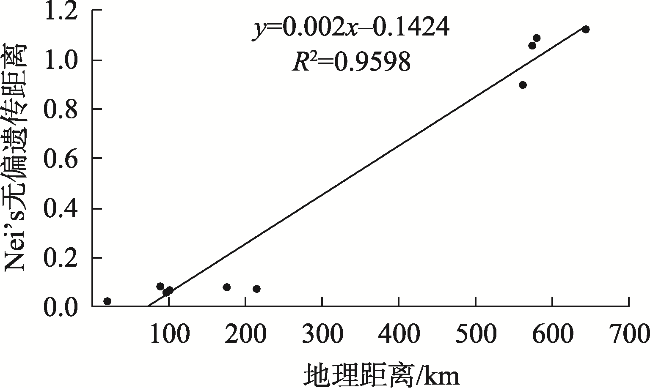
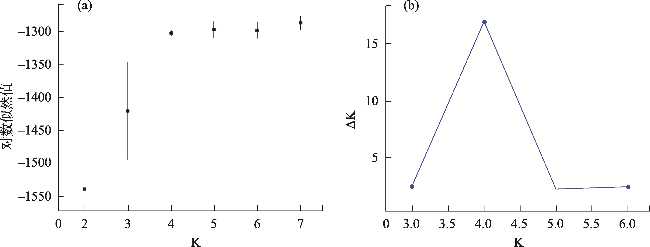
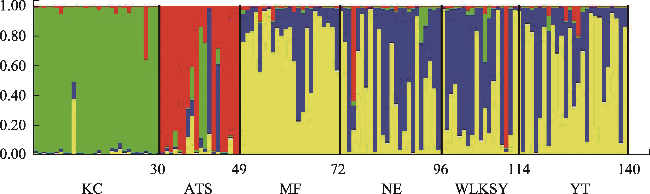
In heterostylous plants, the floral morph composition and frequency of populations are a consequence of mating events over generations. “Homostyly” with anthers and stigmas at the same level within a flower is a floral morph that frequently appears in the evolution of heterostyly and is often accompanied by breakdown of the heteromorphic incompatibility system and the decline of genetic diversity. To explore the formation of the H-morph and its effects on the population, we studied five Limonium aureum populations exhibiting a widely distributed floral morph (H-morph), similar to “homostyly,” in the southern margin of Tarim Basin. The floral morph composition and heterostylous syndrome were investigated by field observations and hand pollination experiments. Meanwhile, the genetic diversity and genetic structure were studied using SSR molecular markers. The results showed the following: (1) The Atushi (ATS) population consisted only of the H-morph, while the remaining four populations were composed of the long- and short-styled morph, and H-morph. All populations had a moderate level of genetic diversity, dimorphism of stigma-pollen morphology, and a strict heteromorphic incompatibility system, revealing that the floral morphs with different pollen ornamentation and stigma papilla cell morphology were compatible. (2) Genetic structure analysis, principal coordinate analysis, and phylogenetic analysis showed that the ATS population had emerged earlier and was independent of the other four populations phylogenetically, with a significant correlation between genetic distance and geographic distance. (3) The H-morphs of the two types of populations may be at different stages of the evolution of heterostyly. The self-incompatibility of the H-morph and the heteromorphic incompatibility system of the population maintained the population’s genetic diversity.
Key words: heterostyly; Limonium aureum; floral variation; genetic diversity; homostyly
表1 12对具有多态性的SSR引物信息Tab. 1 12 pairs of SSR primers’ information with polymorphism |
| 引物名称 | 上游引物序列(5′→3′) | 下游引物序列(5′→3′) | 等位基因大小/bp | 退火温度/℃ |
|---|---|---|---|---|
| LB6 | AGAATCTGAGTGGCTGTT | GGCTTAGGGTTTGTGA | 285~290 | 52 |
| LB12 | CAACTGTGAAAGACGGAAAG | CACGGCAATGGAGGAT | 212~285 | 58 |
| LB26 | AGAGGGTGCCTGGAAA | AGGGTTGATGGCTTGG | 134~140 | 53 |
| LB30 | GTGATAATGCCTGGAGA | GAGGATGTTGGTTTCG | 154~180 | 53 |
| LB64 | CAACTGTGAAAGACGGAAAG | CACGGCAATGGAGGAT | 212~284 | 58 |
| Ld081 | AACCGACGATCCTCTCTTCTC | AGGATTCGCCGGTCTTGT | 144~150 | 65 |
| Ld445 | TCTCTCTCAAGCACCAGCAG | CAGACCGAAGTCAATGAGCA | 196~231 | 60 |
| Ln39 | TGAGCCAATTAGGGCCGCTACCGAG | TCAAGACCCAATGGCTCTGCAGCAACAAAA | 151 | 65 |
| Ln149 | GAGTGGAAATTAACCGACGGA | CTCCATCCCATCATTATTGTACTCATTGTG | 90 | 55 |
| Ln141 | AATTCGATTGCTGCCGAACTG | AATTCATGGACAAGAAGAAGAAGAAGAAGA | 189 | 65 |
| Ln146 | CCCGTTCTTCTCTTCCTCCCTTTG | CCATGGATAGATCCCCGCAATTAGCC | 196 | 62 |
| Ln152 | CAGCACTTTCTATACTAAAACATCGTCGCC | AATTCGCTGGTGAGCCAACCCTATT | 370 | 65 |
表2 11对SSR引物的遗传信息Tab. 2 Genetic informations of 11 pairs of SSR primers |
| 引物名称 | Na | Ne | I | Ho | He | F | Fis | Fst | Nm | PIC |
|---|---|---|---|---|---|---|---|---|---|---|
| LB12 | 5.600 | 2.379 | 1.168 | 0.406 | 0.573 | 0.307 | 0.291 | 0.164 | 1.272 | 0.634 |
| LB26 | 2.200 | 1.234 | 0.333 | 0.125 | 0.184 | 0.194 | 0.318 | 0.489 | 0.262 | 0.270 |
| LB30 | 2.400 | 1.674 | 0.593 | 0.217 | 0.363 | 0.437 | 0.403 | 0.445 | 0.312 | 0.560 |
| LB6 | 3.600 | 1.791 | 0.719 | 0.274 | 0.377 | 0.308 | 0.275 | 0.083 | 2.771 | 0.394 |
| LB64 | 3.600 | 1.816 | 0.737 | 0.330 | 0.408 | 0.158 | 0.190 | 0.281 | 0.640 | 0.470 |
| Ld081 | 2.800 | 2.314 | 0.827 | 0.533 | 0.472 | -0.106 | -0.129 | 0.375 | 0.417 | 0.713 |
| Ld445 | 2.800 | 2.322 | 0.880 | 0.417 | 0.536 | 0.229 | 0.223 | 0.318 | 0.536 | 0.748 |
| Ln141 | 3.000 | 2.392 | 0.919 | 0.672 | 0.554 | -0.209 | -0.213 | 0.162 | 1.296 | 0.590 |
| Ln146 | 2.200 | 1.683 | 0.610 | 0.257 | 0.392 | 0.411 | 0.345 | 0.249 | 0.755 | 0.400 |
| Ln152 | 1.800 | 1.318 | 0.257 | 0.129 | 0.145 | 0.251 | 0.113 | 0.602 | 0.166 | 0.300 |
| Ln39 | 2.400 | 1.673 | 0.379 | 0.187 | 0.188 | -0.020 | 0.005 | 0.344 | 0.477 | 0.255 |
| Mean | 2.945 | 1.872 | 0.675 | 0.322 | 0.381 | 0.178 | 0.166 | 0.319 | 0.809 | 0.485 |
注:Na为观测等位基因数;Ne为有效等位基因数;I为香农信息指数;Ho为观测杂合度;He为期望杂合度;F为固定指数;Fis为近交系数;Fst为居群间分化系数;Nm为基因流;PIC为多态信息含量。下同 |
表3 黄花补血草各居群的遗传信息Tab. 3 Genetic informations of L. aureum populations |
| 居群 | Na | Ne | I | Ho | He | F |
|---|---|---|---|---|---|---|
| ATS | 2.636 | 1.833 | 0.619 | 0.290 | 0.360 | 0.293 |
| MF | 2.273 | 1.581 | 0.514 | 0.232 | 0.304 | 0.203 |
| NE | 3.182 | 2.154 | 0.775 | 0.410 | 0.439 | 0.050 |
| WLKSY | 3.636 | 2.105 | 0.834 | 0.410 | 0.455 | 0.137 |
| YT | 3.000 | 1.688 | 0.630 | 0.269 | 0.346 | 0.211 |
| Mean | 2.945 | 1.872 | 0.675 | 0.322 | 0.381 | 0.178 |
注:ATS为阿图什;MF为民丰县;NE为努尔乡;WLKSY为乌鲁克萨依乡;YT为于田县。下同。 |
表4 5个居群的分子方差分析结果Tab. 4 AMOVA analyse as a weighted average from 5 populations |
| 变异来源 | 自由度 | 均方和 | 方差组分 | 方差分量百分率/% |
|---|---|---|---|---|
| 居群间 | 4 | 82.016 | 0.371 | 12 |
| 居群内 | 215 | 557.6929 | 2.6319 | 88 |
| 总计 | 219 | 693.709 | 3.003 | 100 |
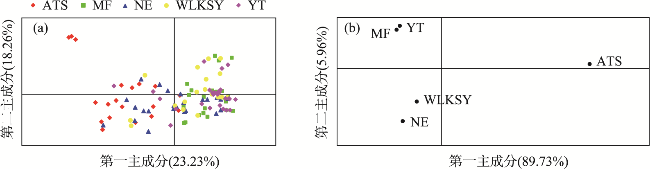
图4 基于遗传距离的5个黄花补血草居群的主坐标分析注:(a)基于个体的分析;(b)基于居群的分析。 Fig. 4 Principal coordinate analysis of 5 L. aureum populations based on genetic distance |
| [1] |
张冬梅, 张华新, 沈熙环, 等. 油松种子园交配系统的时空变化研究[J]. 林业科学, 2004, 40(1): 70-77.
[
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
杨婉舒, 熊昕, 席党鹏, 等. 乌恰县贝壳山: 塔里木盆地西部沧海桑田的见证者[J]. 自然杂志, 2023, 45(2): 148-154.
[
|
| [38] |
潘晓玲. 塔里木盆地植物区系的研究[J]. 新疆大学学报(自然科学版), 1994, 11(4): 77-83.
[
|
| [39] |
欧文雅. 内蒙古境内六种补血草属(Limonium)植物的进化与起源关系初探[D]. 呼和浩特: 内蒙古大学, 2009.
[
|
| [40] |
|
| [41] |
|
| [42] |
阿依古丽·阿卜杜热伊木, 焦芳芳, 张爱勤. 异型花柱植物喀什补血草的传粉者功能群与花粉转移效率[J]. 植物生态学报, 2021, 45(1): 51-61.
[Ayiguli·Abudureyimu,
|
| [43] |
任登芙, 翟雅芯, 张爱勤. 新疆5个驼舌草二型花柱居群交互式雌雄异位的变异[J]. 植物学报, 2023, 58(5): 733-742.
[
|
| [44] |
|
| [45] |
|
| [46] |
刘硕,
[
|
| [47] |
|
| [48] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}