
河西走廊荒漠植物优势种叶片功能性状变异及权衡
|
李善家(1980-),男,教授,博士,主要从事植物逆境生理生态及其分子进化的结构与功能生物学机制. |
收稿日期: 2024-07-08
修回日期: 2024-11-18
网络出版日期: 2025-08-13
基金资助
国家自然科学基金项目(41961007)
中国科学院寒旱区陆面过程与气候变化重点实验室开放基金项目(LPCC2023003)
Variation and trade-offs in leaf functional traits of dominant desert plant species in Hexi Corridor
Received date: 2024-07-08
Revised date: 2024-11-18
Online published: 2025-08-13
李善家 , 李来周 , 缪潆祥 , 苟伟 , 苏培玺 . 河西走廊荒漠植物优势种叶片功能性状变异及权衡[J]. 干旱区研究, 2025 , 42(3) : 480 -488 . DOI: 10.13866/j.azr.2025.03.08
The characteristics of variation in leaf functional traits of different plant types (shrubs, herbaceous plants) in Hexi Corridor and the relationships between these functional traits are extremely important for maintaining the stability of the ecosystem in this region. To explore the mechanisms of adaptation and the ecological strategies of different plant life forms in arid environments, we established survey sites in the eastern, central, and western sections along Hexi Corridor, following a gradient of decreasing natural precipitation from southeast to northwest. We selected 26 dominant desert plant species, including 14 shrubs and 12 herbaceous plants, and analyzed their variability and regional patterns of 14 key leaf functional traits. We also investigated the trade-offs and adaptive strategies among these traits. The following results were obtained: (1) The coefficient of variation for leaf-bound water content (BW), carbon to phosphorus ratio (C:P), plant height (H), and leaf free water content (FW) of dominant desert plant species in the Hexi Corridor exceeds 100%. (2) Plants in different regions of Hexi Corridor displayed diverse survival strategies: plants in the eastern section adopted a “slow-return” strategy, shrubs and herbaceous plants in the central section exhibited “slow-return” and “fast-return” strategies respectively, while shrubs in the western section adopted a “rapid resource acquisition” strategy under resource-rich conditions, and herbaceous plants adopted a “slow-return” strategy under unfavorable soil conditions. The survival strategies of plants are influenced by multiple ecological factors, and they adapt to arid environments through trait optimization and resource allocation.
Key words: desert plants; functional traits; leaf economics spectrum; Hexi Corridor
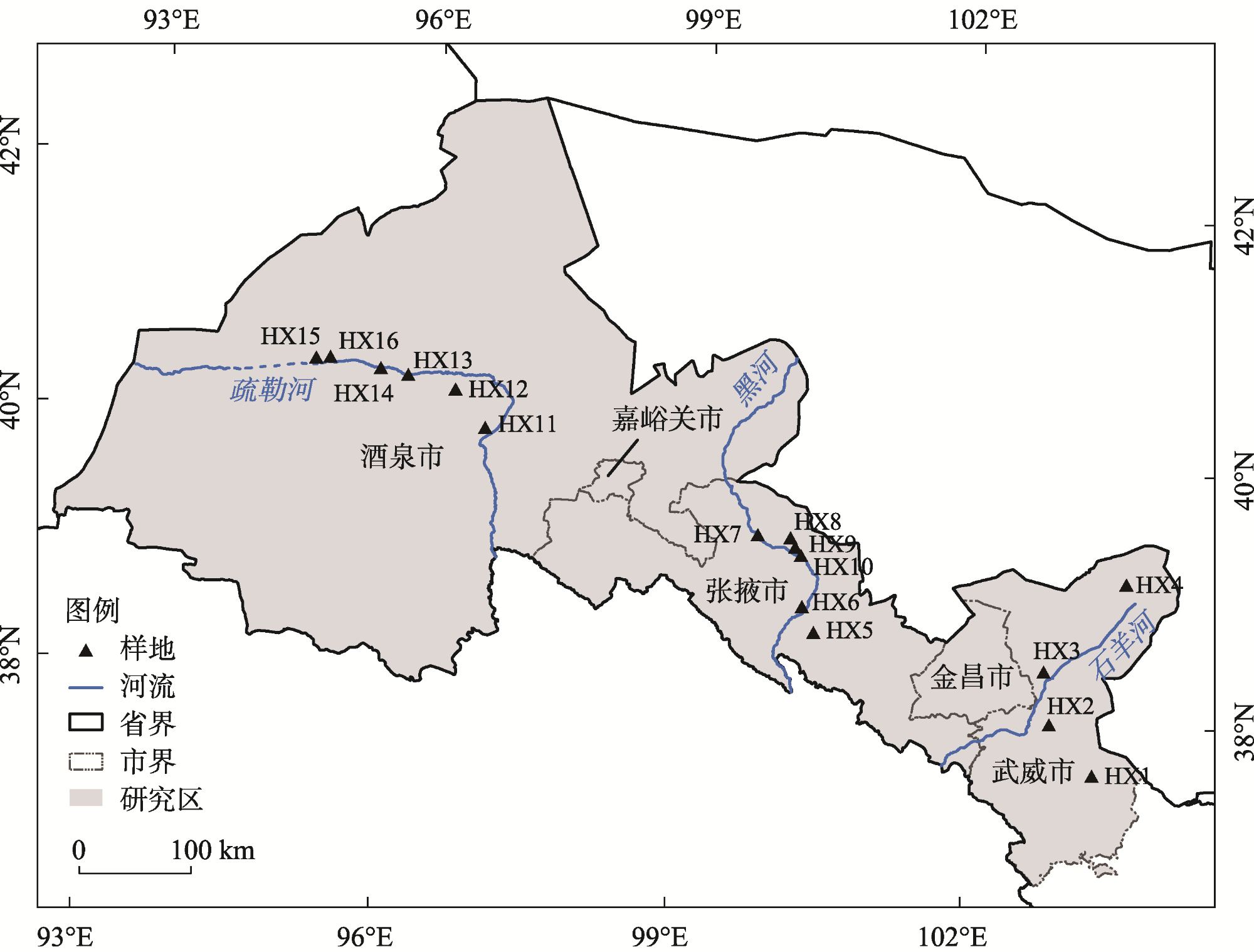
图1 研究区及调查样地分布注: HX1为古浪县黄花滩;HX2为古浪县南湖乡;HX3为民勤县红崖山水库;HX4为民勤县青土湖;HX5为张掖市;HX6为张掖市;HX7为临泽县一工程;HX8为高台县;HX9为临泽县;HX10为临泽县;HX11为瓜州县昌马水库;HX12为瓜州县布隆吉乡;HX13为瓜州县城东;HX14为瓜州县城西;HX15为瓜州县北夹湖;HX16为瓜州县西湖乡北。下同。 Fig. 1 Distribution of study area and survey plots |
表1 河西走廊荒漠植物优势种群落特征Tab. 1 Information of desert plant community in Hexi Corridor sample plot |
| 荒漠区 | 样地编号 | 灌木(重要值) | 草本(重要值) |
|---|---|---|---|
| 东段 | HX1 | 驼绒藜 (0.24)、红砂(0.37) | - |
| HX2 | 沙拐枣(0.54) | 沙米(0.11)、雾冰藜(0.15) | |
| HX3 | 泡泡刺(0.11)、沙拐枣(0.13)、驼绒藜(0.22) | 刺蓬(0.11)、 | |
| HX4 | 白刺(0.18)、梭梭(0.26) | 冰草(0.19) | |
| 中段 | HX5 | 合头草 (0.73) | - |
| HX6 | 红砂(0.33)、珍珠猪毛菜(0.43) | - | |
| HX7 | 红砂(0.21)、泡泡刺(0.24) | - | |
| HX8 | 泡泡刺(0.23)、红砂(0.39) | - | |
| HX9 | 沙拐枣(0.10)、梭梭(0.15) | 雾冰藜(0.19) | |
| HX10 | 梭梭(0.10)、沙拐枣(0.13)、泡泡刺(0.22) | 雾冰藜(0.18) | |
| 西段 | HX11 | 霸王(0.17)、红砂(0.19)、合头草(0.29) | 雾冰藜(0.25) |
| HX12 | 白刺(0.19) | 河西菊(0.77) | |
| HX13 | 黑果枸杞(0.42) | 胀果甘草(0.13)、河西菊(0.23) | |
| HX14 | 黑果枸杞(0.40) | 河西菊(0.26)、芦苇(0.29) | |
| HX15 | 盐爪爪(0.22)、柽柳(0.26)、黑果枸杞(0.37) | - | |
| HX16 | 泡泡刺(0.49) | 芦苇(0.42) |
表2 河西走廊荒漠植物叶性状特征Tab. 2 Leaf traits of desert plants in Hexi Corridor |
| 叶片功能性状 | 灌木 | 草本 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 平均值±标准差 | 变异系数/% | 最大值 | 最小值 | 平均值±标准差 | 变异系数/% | 最大值 | 最小值 | ||
| TWC | 2.94±1.23 | 41.76 | 5.84 | 1.07 | 3.55±2.27 | 63.85 | 9.18 | 0.83 | |
| RWC | 77.72±11.47 | 14.76 | 5.84 | 1.07 | 76.68±14.75 | 19.23 | 98.76 | 45.62 | |
| BW | 1.70±1.19 | 70.3 | 96.17 | 33.9 | 1.20±1.24 | 103.22 | 4.6 | 0.05 | |
| FW | 1.24±0.64 | 51.85 | 4.57 | 0.22 | 2.38±1.56 | 65.42 | 6.18 | 0.6 | |
| BW:FW | 1.96±2.05 | 104.67 | 3.51 | 0.32 | 0.66±0.87 | 132.54 | 4.2 | 0.04 | |
| LDMC | 230.10±70.32 | 30.56 | 9.18 | 0.1 | 228.05±119.48 | 52.39 | 486.8 | 90.8 | |
| SLV | 4.22±1.62 | 38.33 | 392.23 | 128.97 | 4.96±2.11 | 42.58 | 10.13 | 2.22 | |
| SLA | 64.96±19.68 | 30.3 | 8.65 | 1.38 | 81.9±32.55 | 39.74 | 246.86 | 26.11 | |
| LT | 0.83±0.35 | 42.64 | 125.05 | 29.5 | 0.65±0.32 | 48.47 | 1.48 | 0.14 | |
| H | 51.8±41.57 | 80.25 | 200 | 8 | 26.18±18.66 | 71.28 | 77.9 | 6.5 | |
| Suc | 0.06±0.02 | 36.03 | 0.11 | 0.02 | 0.06±0.03 | 45.93 | 0.13 | 0.02 | |
| C | 385.44±71.27 | 18.49 | 1.56 | 0.25 | 366.83±85.15 | 23.21 | 546.27 | 210.1 | |
| N | 22.18±6.98 | 31.49 | 603.4 | 227.6 | 22.52±7.36 | 32.67 | 35.24 | 9.36 | |
| P | 1.37±1.68 | 122.2 | 50.1 | 10.76 | 1.21±0.59 | 48.96 | 2.95 | 0.33 | |
| C:N | 19.40±7.52 | 38.77 | 38.01 | 6.36 | 18.91±10.34 | 54.71 | 58.36 | 9.37 | |
| C:P | 555.67±706.42 | 127.13 | 6304.29 | 40.87 | 382.28±202.82 | 53.05 | 1026.06 | 118.63 | |
| N:P | 34.84±60.13 | 172.57 | 545.75 | 1.88 | 25.98±20.51 | 78.96 | 89.27 | 4.58 | |
注:TWC为叶总含水量,RWC为叶相对含水量,BW为叶束缚水含量,FW为叶自由水含量,BW:FW为叶束缚水与叶自由水比值,LDMC为叶干物质含量,SLV为比叶体积,SLA为比叶面积,LT为叶厚度,C为叶碳含量,N为叶氮含量,P为叶磷含量,C:N为叶碳氮比,C:P为叶碳磷比,N:P为叶氮磷比,H为植株高度,Suc为叶肉质化程度。下同。 |
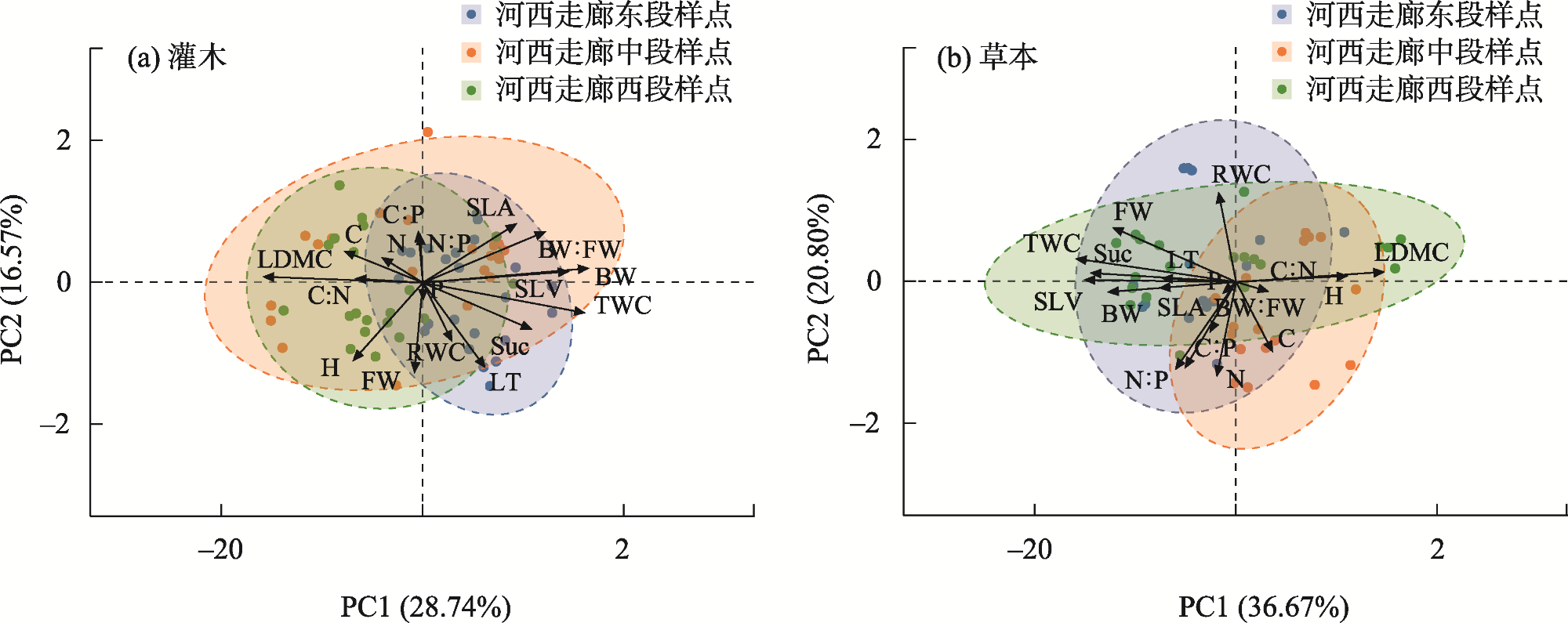
表3 PCA主成分载荷差异Tab. 3 Differences in PCA Principal component loadings |
| 叶片功 能性状 | 灌木 | 草本 | |||
|---|---|---|---|---|---|
| 主成分一 | 主成分二 | 主成分一 | 主成分二 | ||
| TWC | 0.42 | 0.15 | 0.37 | 0.19 | |
| RWC | 0.06 | 0.23 | 0.00 | 0.36 | |
| BW | 0.43 | -0.02 | 0.32 | -0.01 | |
| FW | -0.02 | 0.34 | 0.29 | 0.27 | |
| BW:FW | 0.32 | -0.19 | 0.09 | -0.21 | |
| LDMC | -0.42 | -0.06 | -0.34 | -0.05 | |
| SLV | 0.38 | -0.02 | 0.36 | 0.09 | |
| SLA | 0.20 | -0.32 | 0.15 | 0.03 | |
| LT | 0.16 | 0.42 | 0.15 | 0.24 | |
| C | -0.15 | 0.08 | -0.25 | 0.04 | |
| N | 0.05 | -0.25 | 0.07 | -0.44 | |
| P | 0.04 | 0.19 | -0.15 | 0.28 | |
| H | -0.17 | 0.28 | -0.29 | 0.02 | |
| Suc | 0.27 | 0.33 | 0.36 | 0.13 | |
| N:P | 0.00 | -0.26 | 0.19 | -0.41 | |
| C:N | -0.15 | 0.26 | -0.17 | 0.31 | |
| C:P | 0.00 | -0.25 | 0.14 | -0.31 | |
| 特征值 | 4.88 | 2.82 | 6.23 | 3.55 | |
| 贡献率/% | 28.74 | 16.57 | 36.67 | 20.80 | |
表4 河西走廊东、中、西段灌草叶片功能性状差异比较方差分析结果Tab. 4 ANOVA of the variance of leaf functional traits in Hexi Corridor |
| 叶片功能性状 | 灌木 | 草本 | |||||
|---|---|---|---|---|---|---|---|
| 东段 | 中段 | 西段 | 东段 | 中段 | 西段 | ||
| TWC/g | 2.08b | 2.89a | 3.56a | 2.58ns | 4.28ns | 3.41ns | |
| RWC/% | 76.26ns | 75.95ns | 80.36ns | 77.88a | 67.33b | 86.66a | |
| BW/g | 0.75b | 1.77a | 2.36a | 0.39b | 1.74a | 1.07ab | |
| FW/g | 1.33ns | 1.12ns | 1.20ns | 2.20ns | 2.49ns | 2.34ns | |
| BW:FW | 0.72b | 1.92ab | 3.11a | 0.28ns | 1.64ns | 0.41ns | |
| C/(g·kg-1) | 421.86ns | 377.09ns | 360.27ns | 438.87ns | 305.15ns | 370.15ns | |
| N/(g·kg-1) | 23.83ns | 24.57ns | 20.48ns | 21.78b | 28.61a | 18.59b | |
| P/(g·kg-1) | 1.15ns | 1.02ns | 1.73ns | 1.63ns | 1.09ns | 1.04ns | |
| SLV/(cm3·g-1) | 3.39b | 4.37ab | 4.70a | 4.05ns | 5.50ns | 4.91ns | |
| SLA/(cm2·g-1) | 60.89ns | 65.60ns | 69.42ns | 91.77ns | 90.19ns | 69.39ns | |
| LDMC/(mg·g-1) | 271.08a | 237.53ab | 196.58b | 258.71a | 143.89b | 287.86a | |
| LT/mm | 0.79ab | 0.62b | 0.97a | 0.50ns | 0.61ns | 0.74ns | |
| Suc/(g·g-1) | 0.05b | 0.06ab | 0.07a | 0.04ns | 0.07ns | 0.06ns | |
| H/cm | 80.08a | 36.71b | 40.77b | 28.99ns | 17.23ns | 31.57ns | |
| N:P | 31.54ns | 111.46ns | 27.48ns | 15.31b | 38.95a | 20.57b | |
| C:N | 18.86ns | 17.64ns | 18.73ns | 23.87a | 10.73b | 20.17a | |
| C:P | 407.5ns | 1386.4ns | 454.43ns | 292.22ns | 416.07ns | 388.31ns | |
注:不同字母代表存在显著性差异(P<0.05),ns表示没有显著差异。 |
| [1] |
|
| [2] |
刘晓娟, 马克平. 植物功能性状研究进展[J]. 中国科学: 生命科学, 2015, 45(4): 325-339.
[
|
| [3] |
李耀琪, 王志恒. 植物叶片形态的生态功能、地理分布与成因[J]. 植物生态学报, 2021, 45(10): 1154-1172.
[
|
| [4] |
胡焕琼, 李利, 于军, 等. 四翅滨藜和多枝柽柳对土壤干旱的响应差异[J]. 干旱区研究, 2023, 40(12): 2007-2015.
[
|
| [5] |
余洋, 张志浩, 杨建明, 等. 疏叶骆驼刺叶、根生态化学计量特征对水氮添加的响应[J]. 干旱区研究, 2022, 39(2): 551-559.
[
|
| [6] |
何芸雨, 郭水良, 王喆. 植物功能性状权衡关系的研究进展[J]. 植物生态学报, 2019, 43(12): 1021-1035.
[
|
| [7] |
李善家, 苏培玺, 张海娜, 等. 荒漠植物叶片水分和功能性状特征及其相互关系[J]. 植物生理学报, 2013, 49(2): 153-160.
[
|
| [8] |
马俊梅, 马剑平, 满多清, 等. 河西走廊天然胡杨林的分布和更新特征及其与土壤因子的关系[J]. 干旱区研究, 2023, 40(2): 224-234.
[
|
| [9] |
李敏, 孙杰, 陈雪, 等. 荒漠植物叶片-土壤化学计量及植物内稳态特征[J]. 干旱区研究, 2024, 41(1): 104-113.
[
|
| [10] |
董雪, 李永华, 辛智鸣. 河西走廊西段戈壁灌木群落多样性及其分布格局研究[J]. 干旱区地理, 2020, 43(6): 1514-1522.
[
|
| [11] |
田玉清, 石道良, 张淑倩, 等. 河西走廊水生植物多样性格局、群落特征及影响因素[J]. 生态学报, 2020, 40(1): 202-212.
[
|
| [12] |
孙启兴, 杨晓东, 李浡睿, 等. 水力性状对荒漠植物群落物种多度分布格局的影响[J]. 干旱区研究, 2023, 40(3): 412-424.
[
|
| [13] |
尚佳州, 赵瑜琦, 王卫锋, 等. 干旱对碧玉杨幼苗水氮利用与同化物分配的影响[J]. 干旱区研究, 2022, 39(3): 893-899.
[
|
| [14] |
|
| [15] |
李善家, 王子濠, 苏培玺, 等. 荒漠植物性状权衡策略及功能多样性研究进展[J]. 生态学报, 2022, 42(18): 7308-7320.
[
|
| [16] |
赵文智, 任珩, 杜军, 等. 河西走廊绿洲生态建设和农业发展的若干思考与建议[J]. 中国科学院院刊, 2023, 38(3): 424-434.
[
|
| [17] |
满多清, 李得禄, 刘明成, 等. 民勤西沙窝沙区不同演替阶段植被变化特征研究[J]. 干旱区研究, 2023, 40(12): 1949-1958.
[
|
| [18] |
|
| [19] |
高永龙, 孙艳丽, 徐铭泽, 等. 北京百花山落叶阔叶林群落内木本植物的叶片功能性状变异特征[J]. 北京林业大学学报, 2024, 46(4): 40-51.
[
|
| [20] |
阮成江, 李代琼. 黄土丘陵区沙棘林几个水分生理生态特征研究[J]. 林业科学研究, 2002, 15(1): 47-53.
[
|
| [21] |
戚德辉, 温仲明, 王红霞, 等. 黄土丘陵区不同功能群植物碳氮磷生态化学计量特征及其对微地形的响应[J]. 生态学报, 2016, 36(20): 6420-6430.
[
|
| [22] |
刘玉祯, 刘文亭, 杨晓霞, 等. 放牧对全球草地生态系统碳氮磷化学计量特征影响的Meta分析[J]. 应用生态学报, 2022, 33(5): 1251-1259.
[
|
| [23] |
|
| [24] |
|
| [25] |
李瑞, 单立山, 解婷婷, 等. 典型荒漠灌木叶片功能性状特征随降水梯度的变化研究[J]. 干旱区研究, 2023, 40(3): 425-435.
[
|
| [26] |
|
| [27] |
|
| [28] |
李佳婧, 梁咏亮, 李静尧, 等. 基于叶片功能性状的贺兰山西坡植物生态策略分析[J]. 生态环境学报, 2024, 33(1): 45-53.
[
|
| [29] |
周志宇, 张莉丽, 高文星, 等. 试论灌木是干旱、半干旱区草地恢复中重要的生物资源[J]. 草业科学, 2007, 24(12): 19-21.
[
|
| [30] |
王子婷, 杨磊, 李广, 等. 半干旱黄土区苜蓿退化对坡面草本植物分布及多样性的影响[J]. 生态学报, 2019, 39(10): 3720-3729.
[
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
陈莹婷, 许振柱. 植物叶经济谱的研究进展[J]. 植物生态学报, 2014, 38(10): 1135-1153.
[
|
| [37] |
张姗姗, 张兴, 曲彦婷, 等. 留园植物叶性状及其叶经济谱研究[J]. 北方园艺, 2022(14): 57-65.
[
|
| [38] |
宋贺, 于鸿莹, 陈莹婷, 等. 北京植物园不同功能型植物叶经济谱[J]. 应用生态学报, 2016, 27(6): 1861-1869.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}