
土壤微生物群落对放牧的响应及其与环境因子的关系
|
江康威(1998-),男,硕士研究生,主要研究方向为草地生态恢复. E-mail: waff981021@163.com |
收稿日期: 2024-09-24
修回日期: 2024-11-20
网络出版日期: 2025-08-13
基金资助
国家自然科学基金项目(2522GZRJJ)
国家自然科学基金项目(31960338)
国家自然科学基金项目(41561103)
东北师范大学植被生态科学教育部重点实验室开放项目(130026533)
Responses of soil microbial communities to grazing and their relationship with environmental factors
Received date: 2024-09-24
Revised date: 2024-11-20
Online published: 2025-08-13
江康威 , 王亚菲 , 刘晨通 , 李宏 , 吕程 , 吐尔逊娜依·热依木 , 张青青 . 土壤微生物群落对放牧的响应及其与环境因子的关系[J]. 干旱区研究, 2025 , 42(3) : 467 -479 . DOI: 10.13866/j.azr.2025.03.07
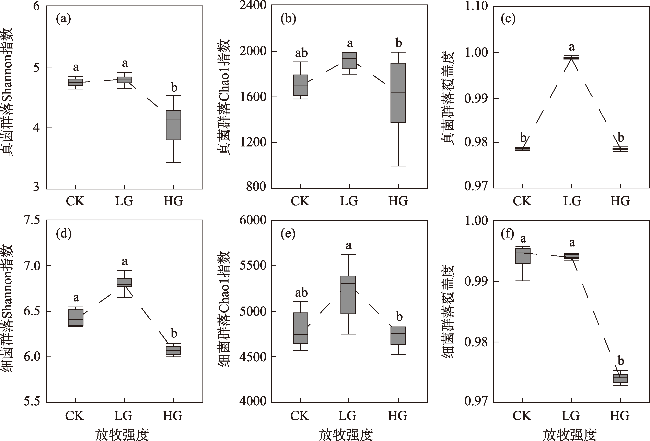
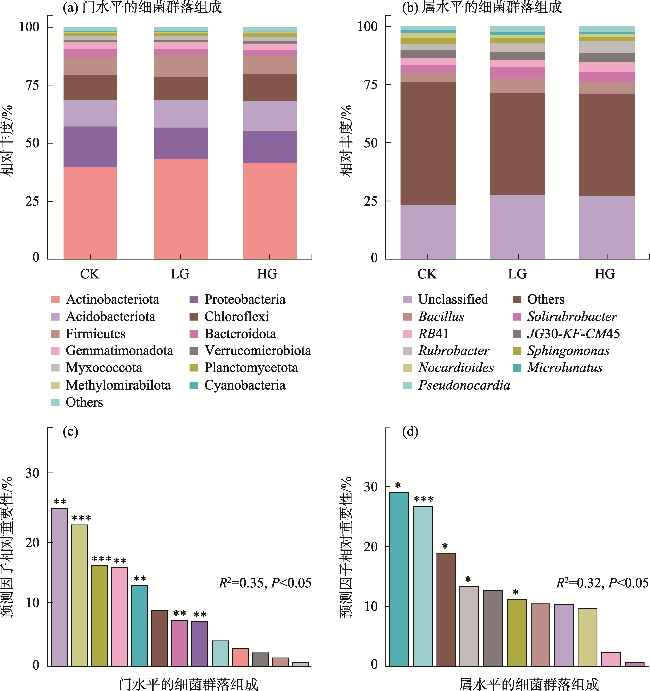
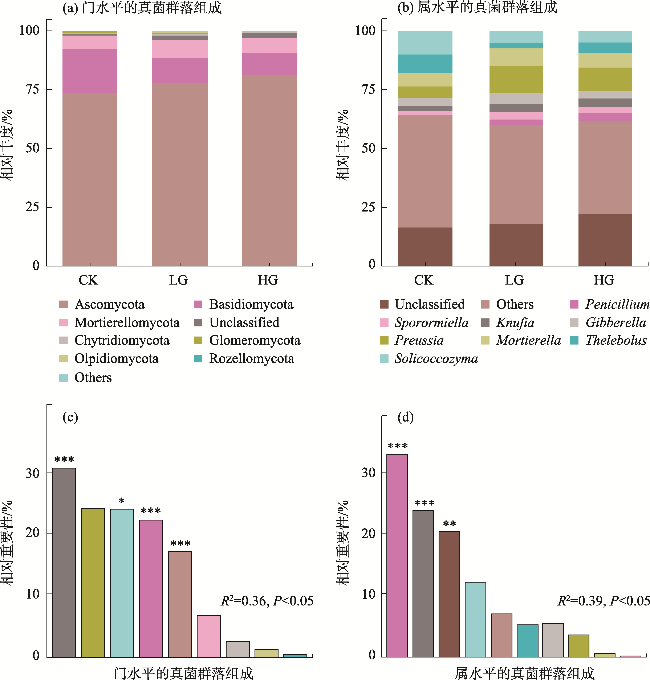
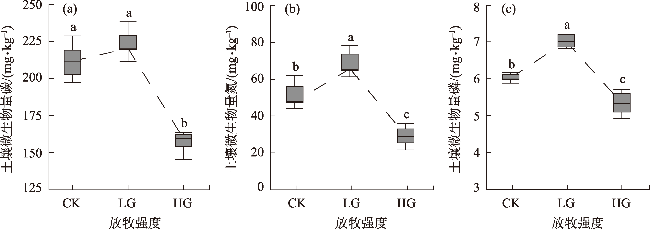
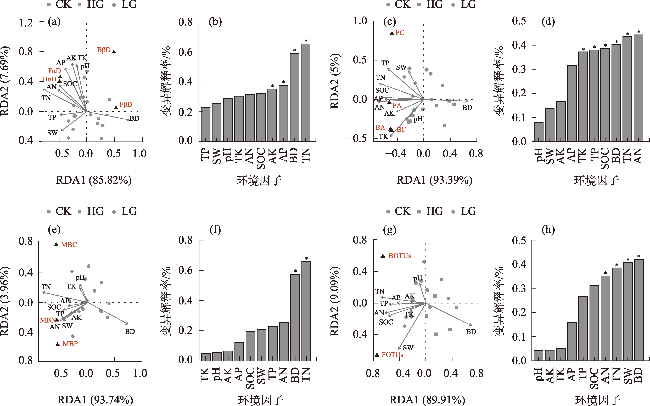
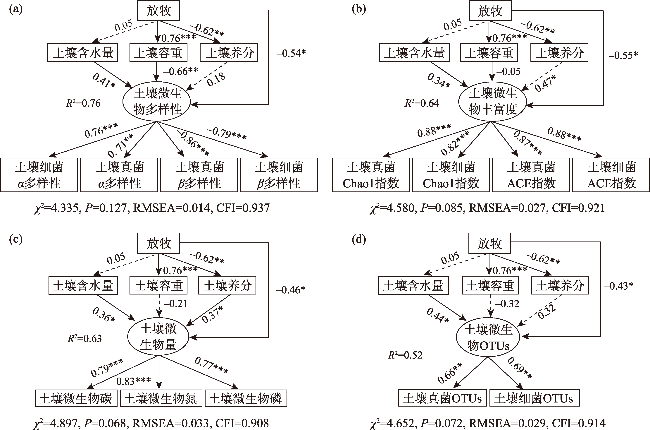
To explore the differences in the responses of different soil microbial community characteristics to grazing intensity, grasslands with different grazing intensities in the middle section of the northern slopes of Tianshan Mountains were selected as a focus for this study. Combining field investigation and indoor analysis, the changing patterns of soil microbial community characteristics with grazing intensity and their intrinsic correlation with soil factors were analyzed. The results showed that Actinobacteria and Ascomycota were the dominant phyla of bacteria and fungi, respectively. Compared with the findings upon heavy grazing, light grazing significantly increased the alpha diversity of microbial communities (P<0.05) and promoted the accumulation of soil microbial biomass carbon, nitrogen, and phosphorus contents. Redundancy analysis and Mantel test showed that the soil microbial community characteristics were positively correlated with the soil total nitrogen and negatively correlated with the soil bulk density (P<0.05). Furthermore, the structural equation model showed that grazing negatively impacted the microbial diversity, richness, biomass, and OTUs characteristics by increasing the bulk density and reducing the soil nutrients (P<0.05). Compared with other indicators, soil microbial community diversity was more sensitive to grazing. In summary, light grazing is conducive to improving the microbial community, and reasonable regulation of grazing intensity is a feasible strategy to ensure the stable development of microbial communities.
表1 研究区植物群落基本特征Tab. 1 Basic characteristics of plant communities in the study area |
| 放牧强度 | |||
|---|---|---|---|
| 轻度放牧 | 重度放牧 | 未放牧 | |
| 植物优势种 | 细果薹草+针茅+无芒雀麦 (Carex stenocarpa+Stipa capillata+Bromus inermis) | 醉马草+细果薹草+平车前(Achnatherum inebrians+Carex stenocarpa+Plantago depressa) | 针茅+羊茅+草地早熟禾 (Carex stenocarpa+Festuca ovina+Poa pratensis) |
| 高度/cm | 18.12±2.15b | 11.53±1.68c | 24.83±2.86a |
| 盖度/% | 92±5.23a | 63±3.76b | 84±3.51a |
| 生物量/(g·m-2) | 186.75±16.73b | 78.24±6.91c | 223.89±14.36a |
| 土壤有机碳/(g·kg-1) | 94.40±2.92a | 91.04±3.39a | 64.05±2.63b |
| 土壤全氮/(g·kg-1) | 2.56±0.55ab | 2.96±0.49a | 1.35±0.46b |
| 全磷/(g·kg-1) | 0.85±0.08a | 0.88±0.06a | 0.71±0.03b |
| 全钾/(g·kg-1) | 10.93±0.57a | 9.10±0.43ab | 8.15±0.42b |
| 速效氮/(mg·kg-1) | 118.46±22.79ab | 129.24±22.91a | 86.04±21.70b |
| 速效磷/(mg·kg-1) | 25.60±1.36a | 20.29±2.23ab | 13.91±1.27b |
| 速效钾/(mg·kg-1) | 438.78±43.11a | 356.49±48.51ab | 304.18±45.99b |
| 土壤密度/(g·cm-3) | 0.65±0.03b | 0.66±0.01b | 0.82±0.04a |
| 土壤含水量/% | 0.47±0.04ab | 0.60±0.03a | 0.41±0.02b |
| 土壤pH | 7.71±0.08a | 7.55±0.15a | 7.15±0.14a |
注:表中数值为平均值±标准误;不同小写字母表示差异性显著。 |
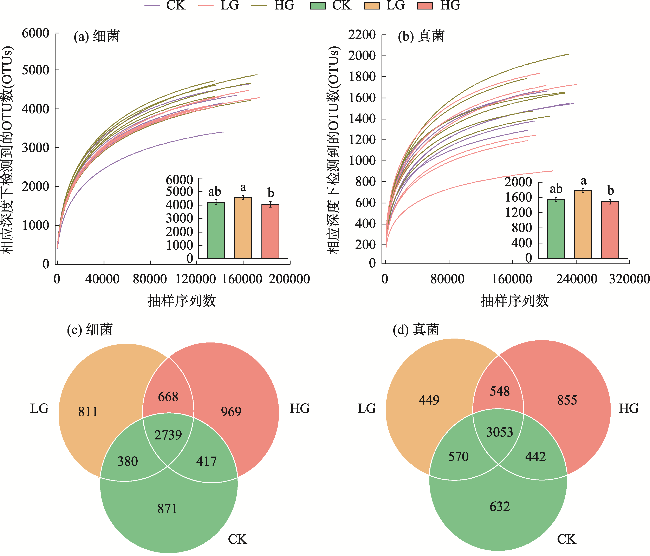
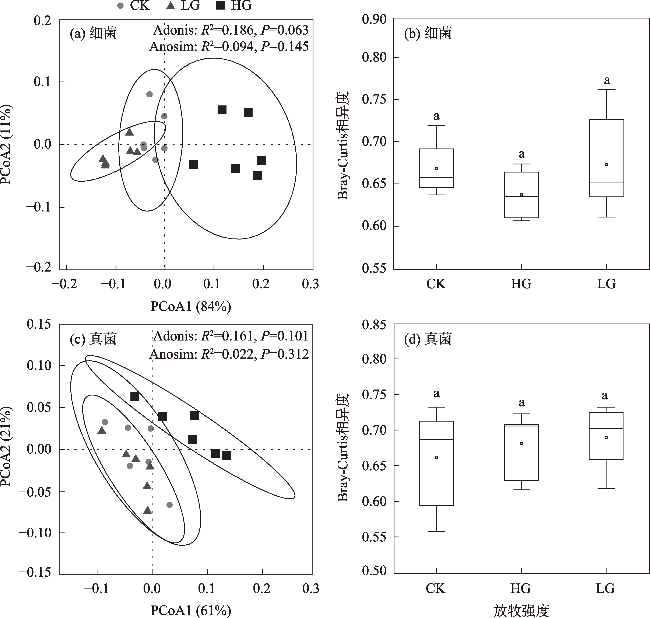
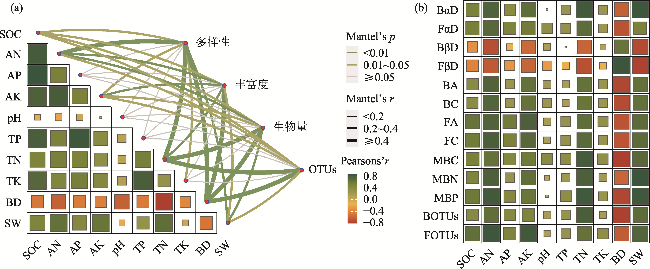
图8 放牧干扰下土壤微生物群落多样性、丰富度、生物量和OTUs与土壤理化因子的冗余分析注:SOC为有机碳;AN为速效氮;AP为速效磷;AK为速效钾;TN为全氮;TP为全磷;TK为全钾;pH为酸碱度;SW为含水量;BD为容重;FαD为真菌α多样性;BαD为细菌α多样性;FβD为真菌β多样性;BβD为细菌β多样性;FA为真菌ACE指数;BA为细菌ACE指数;FC为真菌Chao1指数;BC为细菌Chao1指数;MBC为微生物量碳;MBN为微生物量氮;MBP为微生物量磷;BOTUs为细菌OTU数目;FOTUs为真菌OTU数目。下同。 Fig. 8 Redundancy analysis of soil microbial community diversity, richness, biomass, OTUs and soil physicochemical factors under grazing disturbance |
| [1] |
|
| [2] |
|
| [3] |
郑慧, 薛江博, 桂建华, 等. 放牧强度对华北农牧交错带典型草地土壤化学计量特征的短期影响[J]. 应用生态学报, 2021, 32(7): 2433-2439.
[
|
| [4] |
赵轻舟, 王艳芬, 崔骁勇, 等. 草地土壤微生物多样性影响因素研究进展[J]. 生态科学, 2018, 37(3): 204-212.
[
|
| [5] |
|
| [6] |
|
| [7] |
郑佳华, 赵萌莉, 王琪, 等. 放牧和刈割对大针茅草原土壤微生物群落结构及多样性的影响[J]. 生态学报, 2022, 42(12): 4998-5008.
[
|
| [8] |
张彤, 刘静, 韩叙, 等. 放牧对沙地樟子松林土壤养分及微生物群落的影响[J]. 干旱区研究, 2023, 40(2): 194-202.
[
|
| [9] |
|
| [10] |
苏比努尔·吾麦尔江, 吐尔逊娜依·热依木, 于昭文, 等. 山地草甸草地植物与昆虫多样性对放牧强度的响应[J]. 中国草地学报, 2023, 45(3): 20-29.
[
|
| [11] |
江康威, 张青青, 王亚菲, 等. 放牧干扰下天山北坡中段植物功能群特征及其与土壤环境因子的关系[J]. 植物生态学报, 2024, 48(6): 701-718.
[
|
| [12] |
鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2005.
[
|
| [13] |
|
| [14] |
江康威, 张青青, 王亚菲, 等. 放牧对天山荒漠草地土壤细菌群落的影响[J]. 草业科学, 2023, 40(5): 1243-1257.
[
|
| [15] |
赵文, 尹亚丽, 李世雄, 等. 祁连山不同类型草地土壤细菌群落特征研究[J]. 草业学报, 2021, 30(12): 161-171.
[
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
祁正超, 常佩静, 李永善, 等. 放牧对荒漠灌丛草地土壤团聚体组成及其稳定性的影响[J]. 干旱区研究, 2021, 38(1): 87-94.
[
|
| [20] |
杨阳, 贾丽欣, 乔荠瑢, 等. 重度放牧对荒漠草原土壤养分及微生物多样性的影响[J]. 中国草地学报, 2019, 41(4): 72-79.
[
|
| [21] |
童永尚, 张春平, 董全民, 等. 不同形态氮添加对多年生高寒栽培草地土壤理化性质和微生物群落结构的影响[J]. 环境科学, 2024, 45(6): 3595-3604.
[
|
| [22] |
|
| [23] |
|
| [24] |
王新, 王云英, 裴薇薇, 等. Meta分析放牧对中国草地土壤氮素矿化和硝化作用的影响[J]. 草地学报, 2023, 31(8): 2490-2495.
[
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}