
古尔班通古特沙漠不同坡位地衣结皮土壤磷组分分布特征
|
杨孜悦(2000-),女,硕士研究生,主要从事干旱区生态适应与功能过程研究. E-mail: yangziyuee@163.com |
收稿日期: 2024-12-09
修回日期: 2025-04-02
网络出版日期: 2025-08-13
基金资助
新疆维吾尔自治区天山英才领军(2022TSYCLJ0058)
新疆维吾尔自治区自然科学基金项目(2022D01D83)
新疆维吾尔自治区自然科学基金面上项目(2022D01A349)
新疆维吾尔自治区天池英才
Distribution of soil phosphorus fractions in lichen crusts at different slope positions in Gurbantunggut Desert
Received date: 2024-12-09
Revised date: 2025-04-02
Online published: 2025-08-13
杨孜悦 , 尹本丰 , 张署军 , 黄韵杰 , 杨傲 , 张元明 , 高英志 , 井长青 . 古尔班通古特沙漠不同坡位地衣结皮土壤磷组分分布特征[J]. 干旱区研究, 2025 , 42(7) : 1236 -1245 . DOI: 10.13866/j.azr.2025.07.07
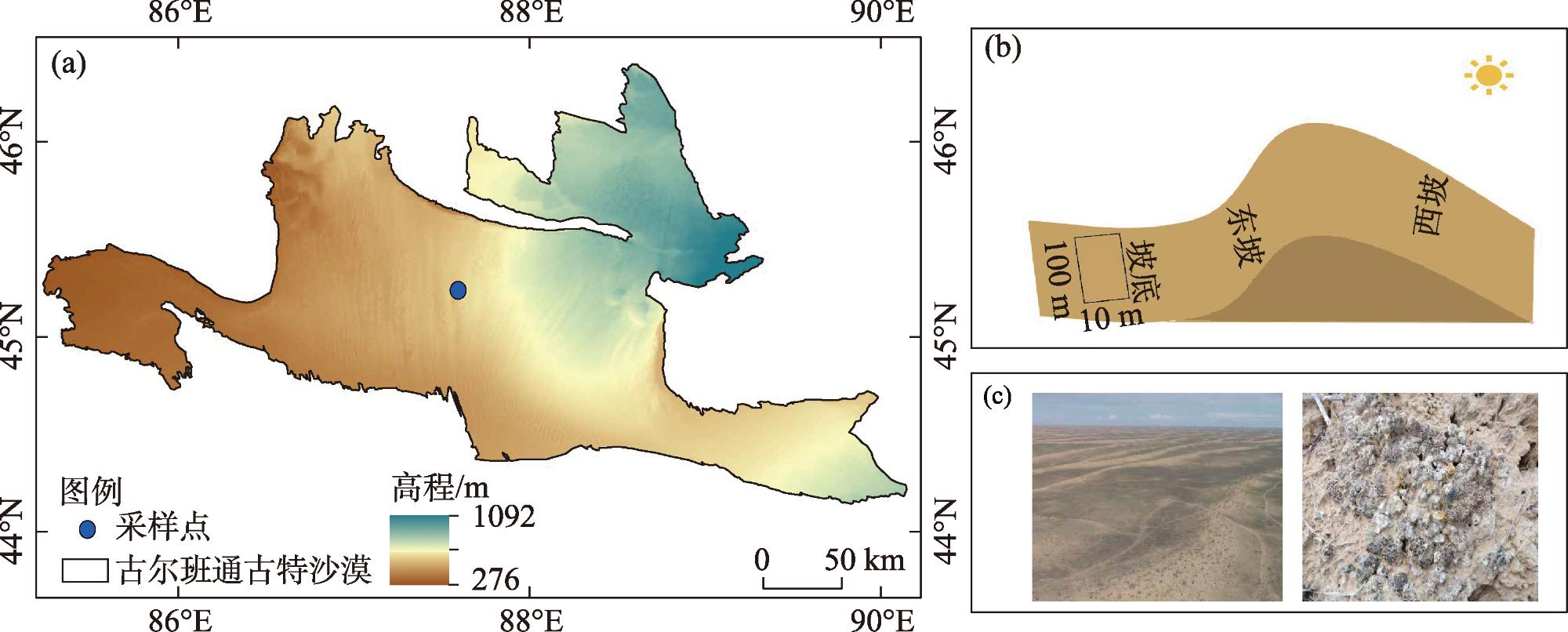
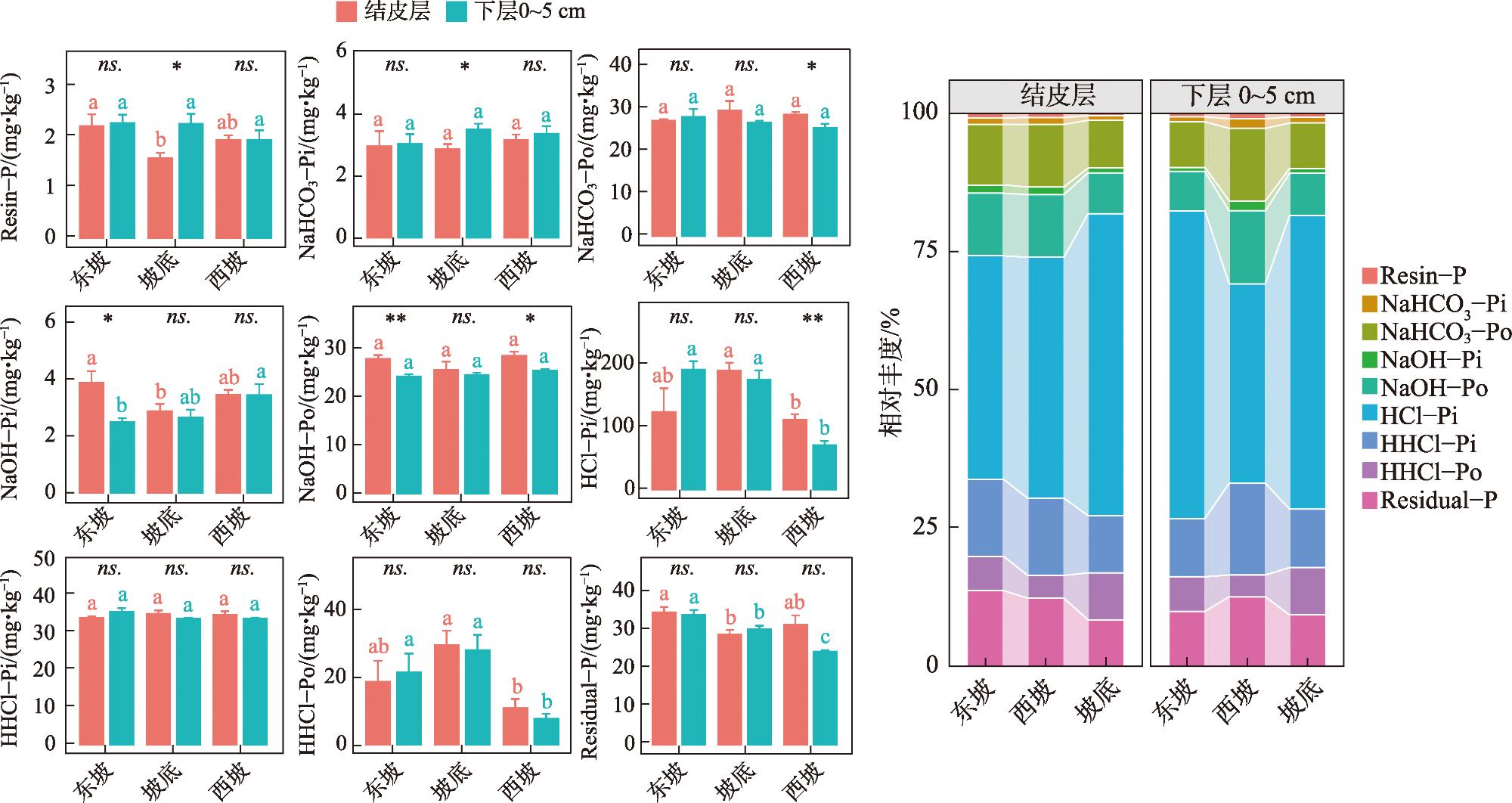
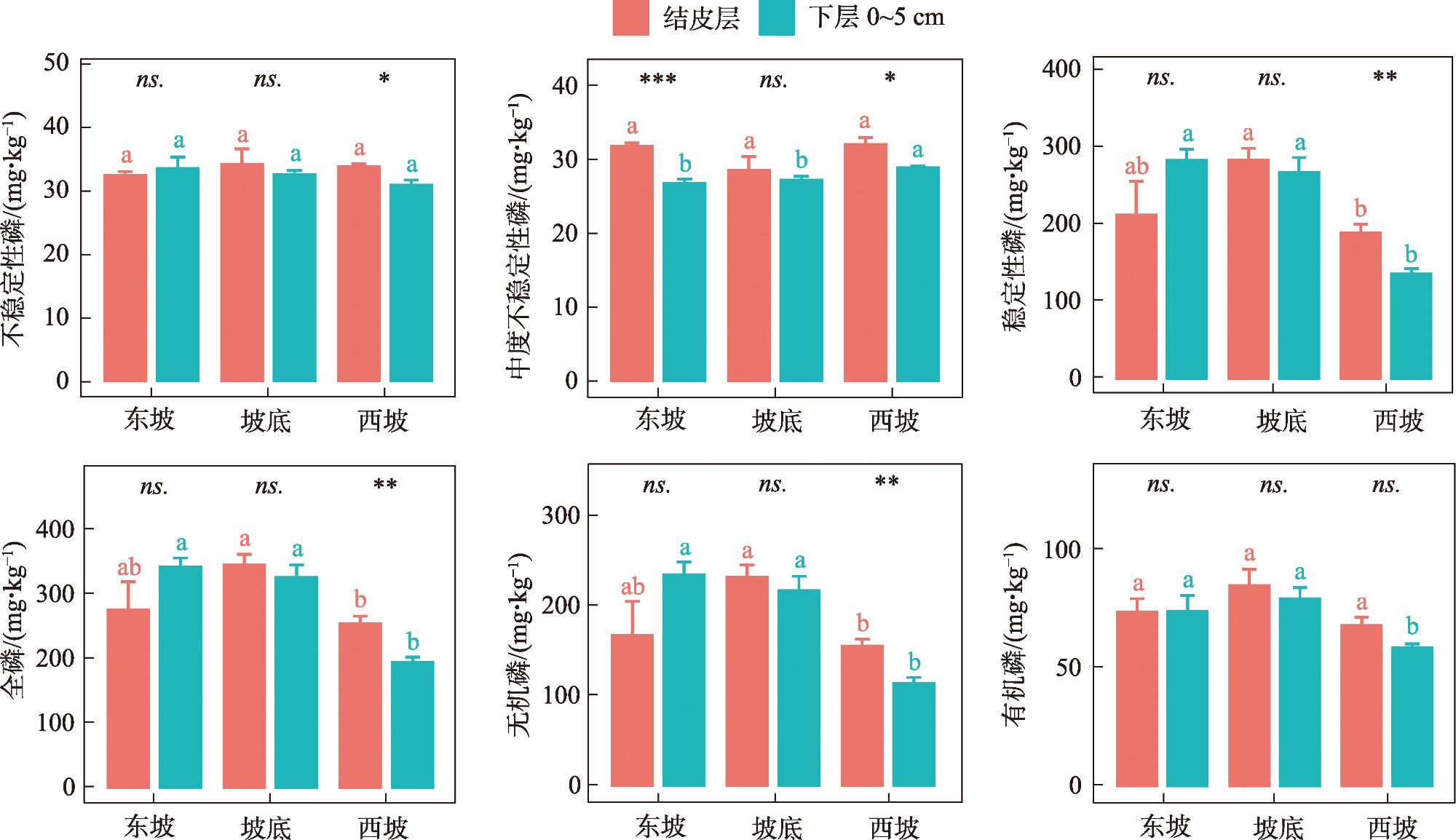
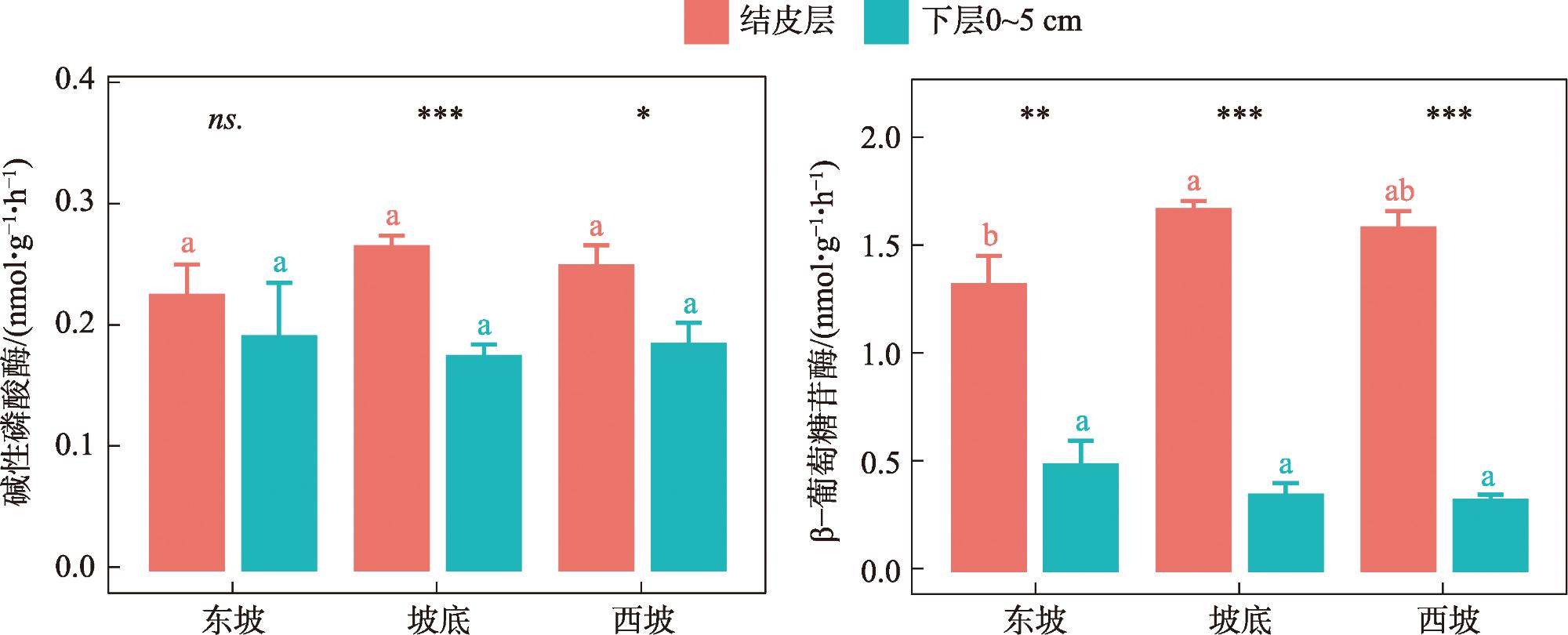
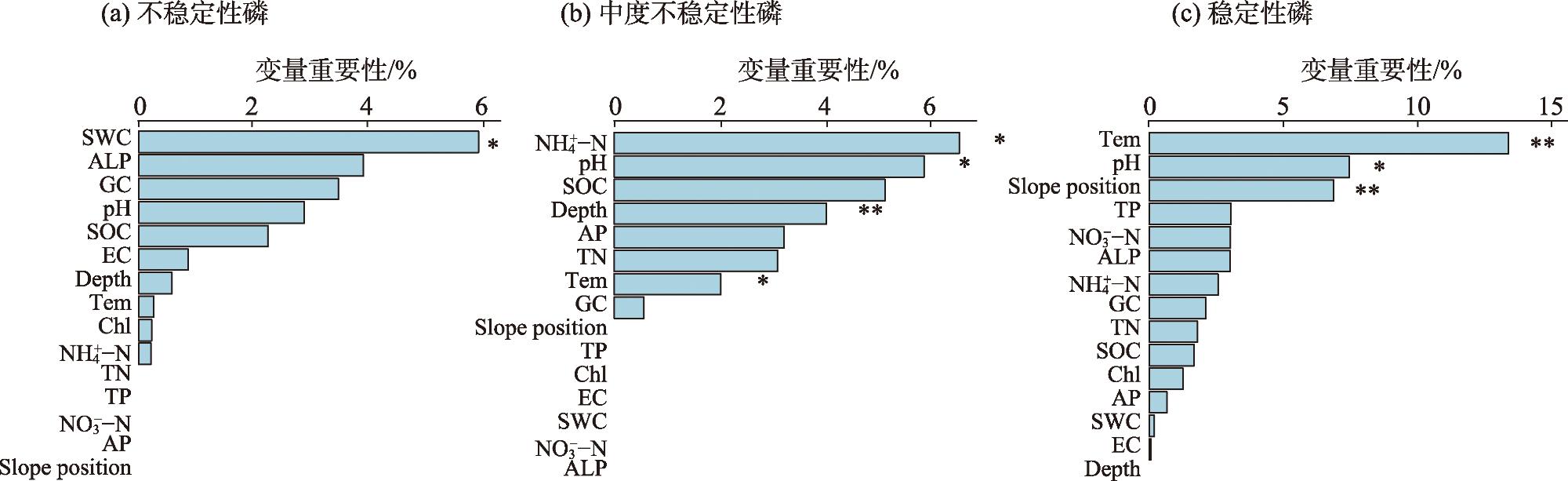
As the fundamental terrain of deserts, sand ridges play a crucial role in shaping the surface water and thermal environment at different slope positions, which profoundly influences how biological soil crusts develop and their spatial distribution patterns. Lichen crusts are widely distributed on the desert surface. However, issues such as how lichen crusts at different slope positions affect the soil phosphorous cycle and what factors play key roles in influencing this remain unclear. Against this background, this study was conducted in Gurbantunggut Desert, involving a systematic analysis of the changes in phosphorus fractions and related enzyme activities in the lichen crust and 0-5 cm soil layer beneath the crust at different slope positions. The results showed that stable phosphorus in the soil (HCl-Pi, HHCl-Po, HHCl-Pi, and Residual-P) accounted for over 75% of the total phosphorus (TP) content, followed by medium labile phosphorus (NaOH-Pi and NaOH-Po) and labile phosphorus (Resin-P, NaHCO3-Pi, and NaHCO3-Po). The slope position had a significant impact on stable phosphorus, and the soil layer had a significant impact on medium labile phosphorus (P<0.05). The data on the contents of stable phosphorus, TP, organic phosphorus (Po), and inorganic phosphorus (Pi) all revealed that, in the crust layer, the values at the bottom of the slope were significantly higher than those on the east and west slopes, while in the 0-5 cm soil layer, the values on the west slope were significantly lower than those at the bottom of the slope and on the east slope (P<0.05). However, the content of NaOH-Pi was significantly higher on the east and west slopes than at the bottom of the slope in the crust layer, and it was significantly higher on the west slope than on the east slope and at the bottom of the slope in the 0-5 cm soil layer. In terms of soil enzymes, the east slope exhibited the lowest activity of alkaline phosphatase activity (ALP) and β-glucosidase activity (GC) in the crust layer, but the highest in the 0-5 cm soil layer. Random forest model analysis showed that the changes in moisture and temperature brought about by the slope position were the most important factors affecting the levels of labile phosphorus and stable phosphorus in the crust soil, respectively. This provides scientific support that enriches the theoretical framework of soil phosphorous cycling in desert ecosystems.
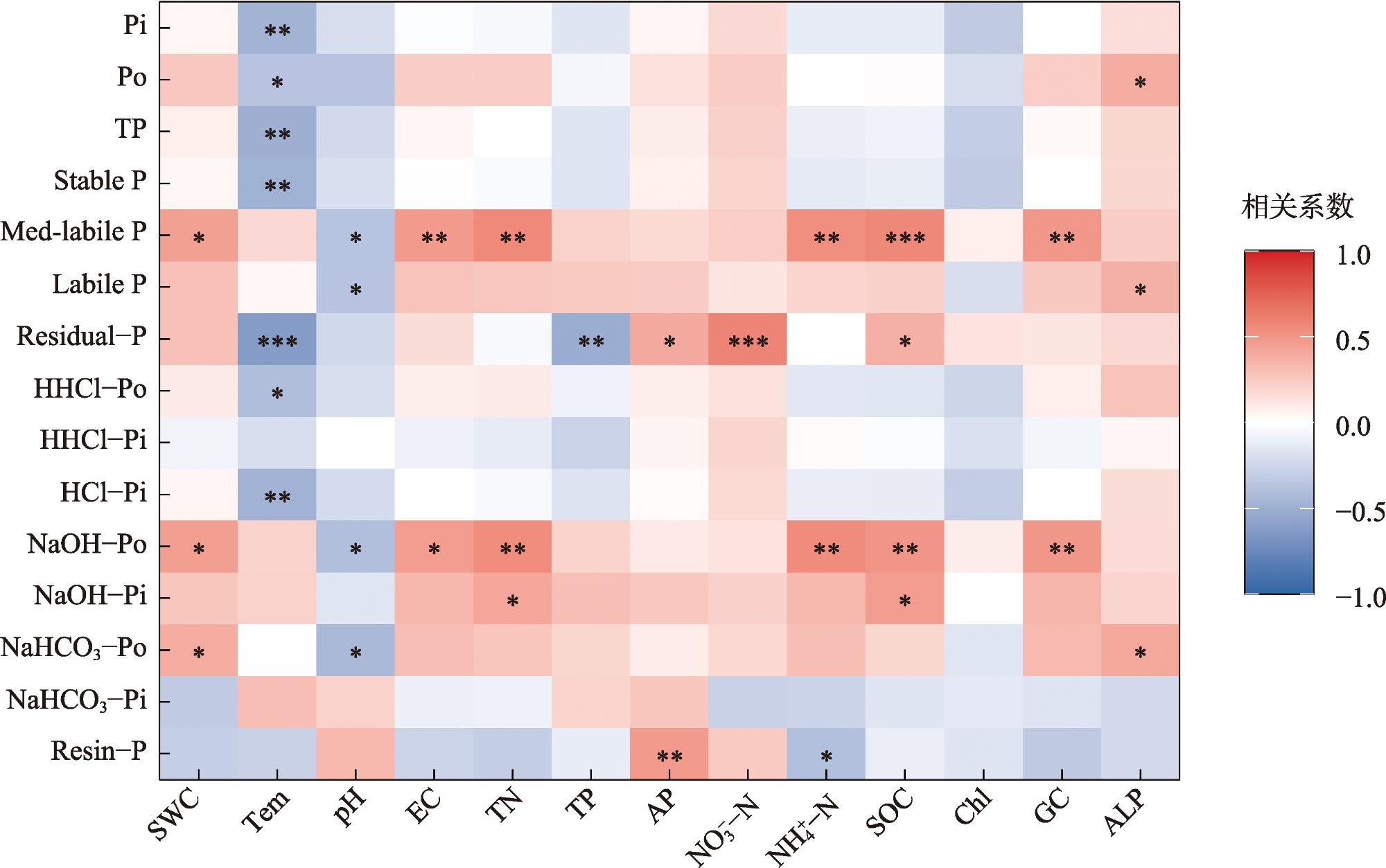
图2 不同坡位对地衣结皮层、下层0~5 cm土壤磷组分含量的影响注:小写字母表示同一土层不同坡位土壤磷组分变化的差异(P<0.05);不同符号表示同一坡位不同土层间的差异。*表示P<0.05;**表示P<0.01;***表示P<0.001;ns.表示不显著。下同。 Fig. 2 Effects of different slope positions on the contents of soil phosphorus fractions in the lichen crust layer and the lower 0-5 cm soil layer |
表1 不同坡位与土层对土壤磷组分影响的双因素方差分析(F值)Tab. 1 Two-way analysis of variance for the effects of different slope positions and soil layers on soil phosphorus fractions (F value) |
| 磷组分与胞外酶 | 土层 | 坡位 | 土层×坡位 |
|---|---|---|---|
| 树脂磷(Resin-P) | 3.151 | 2.309 | 2.413 |
| 碳酸氢钠无机磷(NaHCO3-Pi) | 1.619 | 0.479 | 0.515 |
| 碳酸氢钠有机磷(NaHCO3-Po) | 2.536 | 0.362 | 1.544 |
| 氢氧化钠无机磷(NaOH-Pi) | 5.493* | 2.927 | 3.321 |
| 氢氧化钠有机磷(NaOH-Po) | 12.773** | 2.254 | 1.113 |
| 稀盐酸无机磷(HCl-Pi) | 0.071 | 12.371** | 4.463* |
| 浓盐酸无机磷(HHCl-Pi) | 0.993 | 0.967 | 2.082 |
| 浓盐酸有机磷(HHCl-Po) | 0.029 | 9.852** | 0.242 |
| 残余磷(Residual-P) | 3.226 | 10.856** | 4.614* |
| β-葡萄糖苷酶GC | 264.818** | 0.729 | 4.820* |
| 碱性磷酸酶ALP | 10.133** | 0.134 | 0.675 |
注:*表示P<0.05;**表示P<0.01。 |
| [1] |
|
| [2] |
李承义, 何明珠, 唐亮. 荒漠生态系统磷循环及其驱动机制研究进展[J]. 生态学报, 2022, 42(12): 5115-5124.
[
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
陶冶, 刘耀斌, 吴甘霖, 等. 准噶尔荒漠区域尺度浅层土壤化学计量特征及其空间分布格局[J]. 草业学报, 2016, 25(7): 13-23.
[
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
董鹏, 任悦, 高广磊, 等. 呼伦贝尔沙地樟子松枯落物和土壤碳、氮、磷化学计量特征[J]. 干旱区研究, 2024, 41(8): 1354-1363.
[
|
| [17] |
|
| [18] |
|
| [19] |
雷菲亚, 李小双, 陶冶, 等. 西北干旱区藓类结皮覆盖下土壤多功能性特征及影响因子[J]. 干旱区研究, 2024, 41(5): 812-820.
[
|
| [20] |
张婷婷, 程向敏, 魏鑫丽, 等. 荒漠地衣结皮研究进展[J]. 菌物学报, 2021, 40(1): 1-13.
[
|
| [21] |
张元明, 杨维康, 王雪芹, 等. 生物结皮影响下的土壤有机质分异特征[J]. 生态学报, 2005, 25(12): 3420-3425.
[
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
罗朝逸, 吴艳宏. 高山生态系统生物土壤结皮对磷循环影响研究进展[J]. 土壤通报, 2024, 55(3): 867-875.
[
|
| [36] |
刘春增, 张成兰, 张琳, 等. 长期紫云英配施减量化肥对土壤铵态氮吸附解吸特征的影响[J]. 中国土壤与肥料, 2024(6): 11-17.
[
|
| [37] |
|
| [38] |
|
| [39] |
冯欢欢, 高思齐, 高晋丽, 等. 氮添加对大兴安岭泥炭地典型植物叶片叶绿素和养分含量的影响[J/OL]. 生态学杂志, 1-10[2025-04-22]. http://kns.cnki.net/kcms/detail/21.1148.Q.20240612.1213.004.html.
[
|
| [40] |
|
| [41] |
|
| [42] |
韩志立, 尹本丰, 杨孜悦, 等. 积雪变化对温带荒漠齿肋赤藓结皮土壤磷组分的影响[J]. 生态学报, 2024, 44(16): 7119-7129.
[
|
| [43] |
韩炳宏, 牛得草, 贺磊, 等. 生物土壤结皮发育及其影响因素研究进展[J]. 草业科学, 2017, 34(9): 1793-1801.
[
|
| [44] |
|
| [45] |
|
| [46] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}