
气候变化下菟丝子属3种植物在中国的潜在地理分布
|
瞿宇阳(2000-),男,硕士研究生,研究方向为植物分类学. E-mail: 1071408474@qq.com |
收稿日期: 2024-08-14
修回日期: 2024-10-21
网络出版日期: 2025-08-13
基金资助
国家自然科学基金(31870191)
新疆维吾尔自治区农业外来入侵物种普查(2022-2023东疆专项)
新疆生产建设兵团农业外来入侵物种普查(兵农科发〔2021〕139号)
Potential geographic distribution of the three species of Cuscuta in China under climate scenarios
Received date: 2024-08-14
Revised date: 2024-10-21
Online published: 2025-08-13
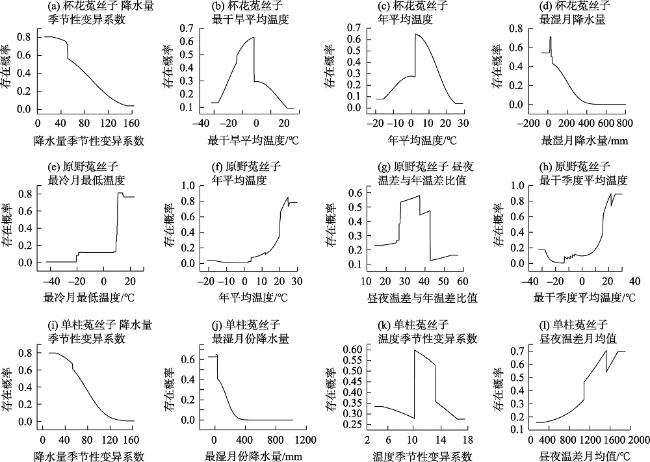
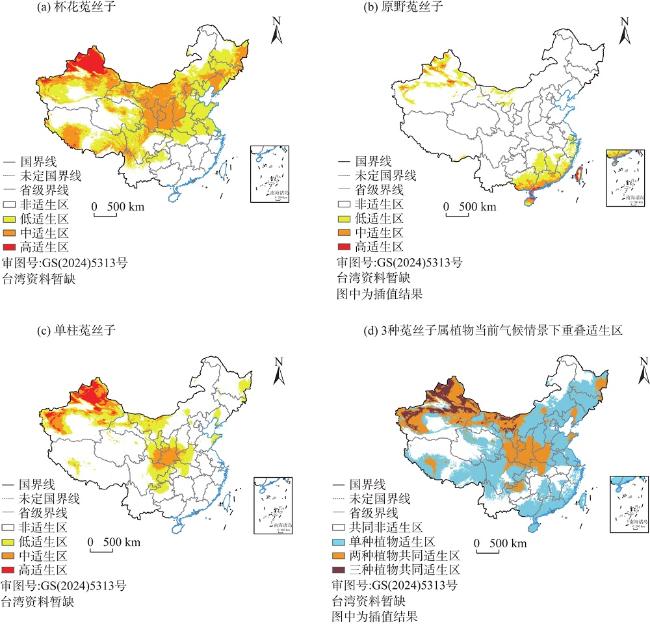
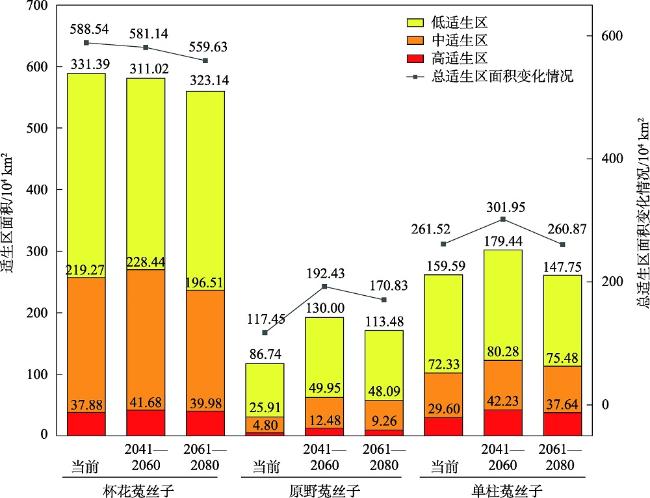
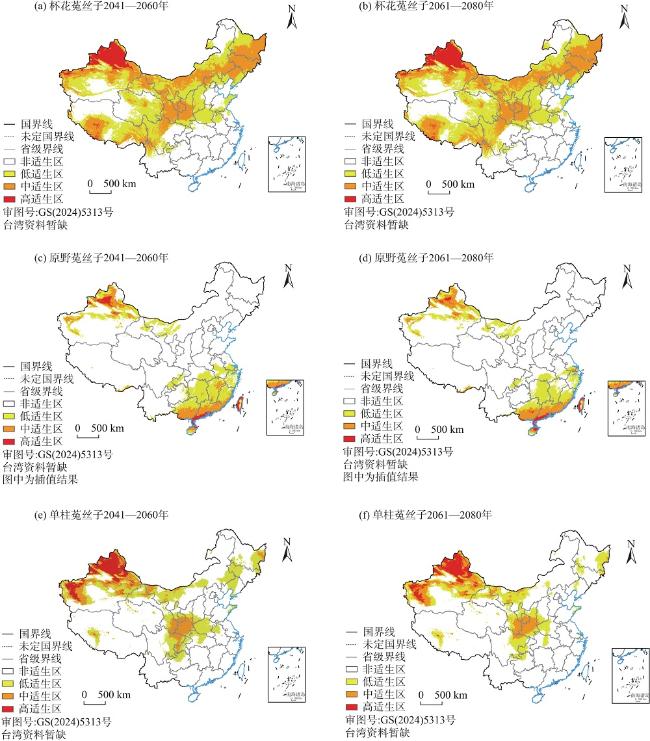
菟丝子属(Cuscuta)植物因茎全寄生的特点全属均被列为检疫植物,探究菟丝子属植物在中国潜在空间分布特点对其防治有重要作用。本研究选取杯花菟丝子(C. cupulata)、原野菟丝子(C. campestris)、单柱菟丝子(C. monogyna)3种菟丝子属植物为研究对象,结合其已有地理分布记录932条、野外考察记录144条和20个环境变量,应用最大熵模型和ArcGIS,预测其在当前和未来(2041—2060年、2061—2080年)SSP2-4.5温室气体排放浓度场景下的潜在适生区分布,并将当前气候情景下菟丝子属3种植物的适生区与土地利用数据进行叠加分析。结果表明:(1) 受试者工作特征曲线下的面积(AUC)均大于0.9,预测精度较高。(2) 影响菟丝子属3种植物适生区分布的主要环境因子存在差异,但均是气候因子占主导,地形因子的贡献率相对较小。(3) 当前气候情景下杯花菟丝子与单柱菟丝子适生区多分布于我国北方,而原野菟丝子适生区除了在北方有分布外亦在东南沿海有大面积覆盖,并且未来3种菟丝子属植物均表现出向我国北部扩张的趋势。(4) 当前时期菟丝子属3种植物的适生区大部分土地已经被开发利用。
瞿宇阳 , 文田田 , 刘佳敏 , 阎平 . 气候变化下菟丝子属3种植物在中国的潜在地理分布[J]. 干旱区研究, 2025 , 42(1) : 97 -107 . DOI: 10.13866/j.azr.2025.01.09
Cuscuta plants are classified as quarantine plants due to their parasitic stem characteristics, and it is important to investigate the potential spatial distribution characteristics of Cuscuta plants in China for effective control. In this study, three species of Cuscuta (C. cupulata, C. campestris and C. monogyna) were selected as the research subjects. Based on 932 records of geographical distribution, 144 records of field investigation and 20 environmental variables, the maximum entropy model and ArcGIS were used to predict potential niche distributions under current and future scenarios (2041-2060, 2061-2080) considering of greenhouse gas emissions from SSP2-4.5. The suitable areas for the 3 species of Cuscuta under the current climate conditions and land use date were analyzed. The results showed that: (1) The area under the work characteristic curve (AUC) for all subjects was greater than 0.9 indicating high prediction accuracy. (2) The main environmental factors affecting the distribution of the three species of Cuscuta within their habitable zones with a relatively small contribution rate from topographic factors varied, but all of them were dominated by climatic factors. (3) Currently C. cupulata and C. monogyna distributions are concentrated in northern China, while C. campestris is not only distributed in the north, but also concentrated in southeastern China, and in future scenarios they will expand further northward. (4) In the current period, most land areas suitable for these three species of Cuscuta have already been exploited.
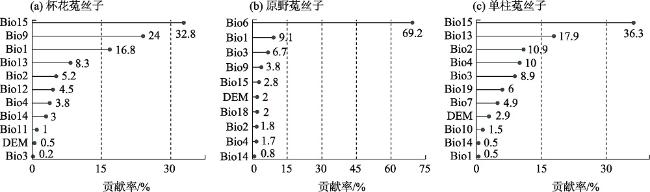
表1 候选环境变量因子Tab. 1 Candidate environment variable factors |
| 环境变量代码 | 变量描述 | 单位 |
|---|---|---|
| Bio1 | 年平均温度 | ℃ |
| Bio2 | 昼夜温差月均值 | ℃ |
| Bio3 | 昼夜温差与年温差比值 | - |
| Bio4 | 温度季节性变化标准差 | - |
| Bio5 | 最热月最高温 | ℃ |
| Bio6 | 最冷月最低温 | ℃ |
| Bio7 | 气温年较差 | ℃ |
| Bio8 | 最湿季平均温度 | ℃ |
| Bio9 | 最干季平均温度 | ℃ |
| Bio10 | 最热季平均温度 | ℃ |
| Bio11 | 最冷季平均温度 | ℃ |
| Bio12 | 年降水量 | mm |
| Bio13 | 最湿月降水量 | mm |
| Bio14 | 最干月降水量 | mm |
| Bio15 | 降水量季节性变异系数 | - |
| Bio16 | 最湿季降水量 | mm |
| Bio17 | 最干季降水量 | mm |
| Bio18 | 最热季降水量 | mm |
| Bio19 | 最冷季降水量 | mm |
| DEM | 海拔 | m |
表2 用于构建模型的环境变量因子Tab. 2 The environment variable factors used to build the model |
| 物种 | 代码 |
|---|---|
| 杯花菟丝子 | Bio1、Bio2、Bio3、Bio4、Bio9、Bio11、Bio12、Bio13、 Bio14、Bio15、DEM |
| 原野菟丝子 | Bio1、Bio2、Bio3、Bio4、Bio6、Bio9、Bio14、Bio15、 Bio18、DEM |
| 单柱菟丝子 | Bio1、Bio2、Bio3、Bio4、Bio7、Bio10、Bio13、Bio15、 Bio19、DEM |
表3 基于kuenm包优化的MaxEnt参数设置Tab. 3 MaxEnt parameter settings optimized based on the kuenm package |
| 类型 | 物种 | 特征组合 | 倍频 | 信息量准则 |
|---|---|---|---|---|
| 默认 | 杯花菟丝子 | LQPH | 1 | 32.93 |
| 原野菟丝子 | LQPH | 1 | 40.83 | |
| 单柱菟丝子 | LQPH | 1 | 82.31 | |
| 优化 | 杯花菟丝子 | QPT | 1.6 | 0 |
| 原野菟丝子 | QPT | 2.8 | 0 | |
| 单柱菟丝子 | QPT | 1.9 | 0 |
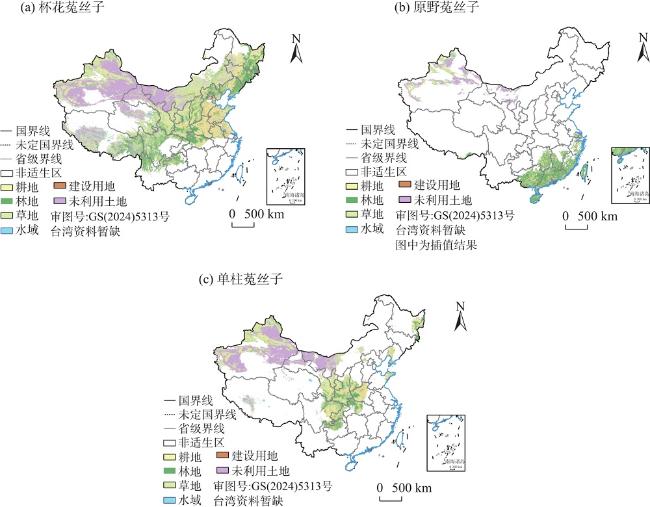
图7 当前气候情景下3种菟丝子属植物潜在适生区土地利用类型分布Fig. 7 Distribution of land types in three potential suitable areas of Cuscuta under current climate scenario |
表 4 当前气候情景下3种菟丝子属植物潜在适生区中土地利用类型分布比例Tab. 4 Distribution proportion of land use types in three potential suitable areas of Cuscuta species under current climate scenarios |
| 土地利用类型 | 耕地/% | 林地/% | 草地/% | 水域/% | 建设用地/% | 未利用土地/% | 总适生区面/104 km2 |
|---|---|---|---|---|---|---|---|
| 杯花菟丝子 | 20.69 | 16.80 | 30.53 | 2.44 | 3.24 | 26.30 | 588.54 |
| 原野菟丝子 | 21.62 | 37.13 | 13.71 | 2.82 | 4.50 | 20.22 | 117.45 |
| 单柱菟丝子 | 21.86 | 13.05 | 24.95 | 2.53 | 2.51 | 35.10 | 261.52 |
| [1] |
周杰, 王旭虎, 杜维波, 等. 气候变化背景下的天山云杉潜在分布区预测[J]. 干旱区研究, 2024, 41(7): 1167-1176.
[
|
| [2] |
王丽丽, 杨采青, 王瑛, 等. 全球入侵物种马铃薯块茎蛾生态位转移及适生区扩展[J]. 应用生态学报, 2024, 35(3): 797-805.
[
|
| [3] |
|
| [4] |
罗瑞萍, 赵志刚, 赵越, 等. 3个主栽大豆品种被菟丝子寄生危害的差异性研究[J]. 农业灾害研究, 2016, 6(1): 3-4.
[
|
| [5] |
尚佰晓. 菟丝子属植物在园林绿化中的危害调查及防治[J]. 园艺与种苗, 2022, 42(8): 28-32.
[
|
| [6] |
周鑫, 瞿巧文, 吴夏华. 菟丝子对林木的危害及其防治[J]. 北方农业学报, 2013(6): 93.
[
|
| [7] |
陈莹, 兰芳, 刘金平, 等. 日本菟丝子对园林植物危害度及适应性生长的研究[J]. 四川林业科技, 2023, 44(1): 32-39.
[
|
| [8] |
|
| [9] |
余波, 瞿添添, 刘应蛟, 等. 菟丝子和大菟丝子形态组织研究与分类学意义[J]. 种子, 2023, 42(10): 57-64.
[
|
| [10] |
|
| [11] |
周冰颖, 刘霞, 谢伟, 等. 我国长江流域园林常见菟丝子及其防治方法[J]. 杂草学报, 2022, 40(3): 8-13.
[
|
| [12] |
陈鲁宁, 胡扬, 辛国松, 等. 菟丝子化学成分、药理作用研究进展及其质量标志物(Q-Marker)预测[J]. 中草药, 2024, 55(15): 5298-5314.
[
|
| [13] |
|
| [14] |
张晓玮, 蒋玉梅, 毕阳, 等. 基于MaxEnt模型的中国沙棘潜在适宜分布区分析[J]. 生态学报, 2022, 42(4): 1420-1428.
[
|
| [15] |
胡永云. 复杂气候系统和全球变暖[J]. 物理, 2022, 51(1): 10-15.
[
|
| [16] |
田叙辰, 魏洪玲, 解胜男, 等. 基于MaxEnt模型的东北地区槭树潜在地理分布[J]. 生态环境学报, 2024, 33(4): 509-519.
[
|
| [17] |
段义忠, 王佳豪, 王驰, 等. 未来气候变化下西北干旱区4种扁桃亚属植物潜在适生区分析[J]. 生态学杂志, 2020, 39(7): 2193-2204.
[
|
| [18] |
|
| [19] |
|
| [20] |
吴帆, 刘国豪, 谢鑫, 等. 未来气候变化对金花茶分布格局的影响[J]. 云南农业大学学报(自然科学), 2024, 39(3): 144-151.
[
|
| [21] |
|
| [22] |
艾尼古丽·依明. 石河子地区菟丝子种类鉴定、生物学特性研究及生防菌筛选[D]. 石河子: 石河子大学, 2017.
[ Ainiguli. Yiming. Species Identification, Biological Characteristics Study, and Screeningof Control Strain of Parasitic Weed Cuscuta in Shihezi[D]. Shihezi: Shihezi University, 2017. ]
|
| [23] |
任子春. 基于Maxent模型在全球变暖条件下菟丝子属全寄生植物及其寄主的潜在分布预测[D]. 太原: 山西师范大学, 2020.
[
|
| [24] |
马金双. 中国入侵植物名录[M]. 北京: 高等教育出版社, 2013: 131-133.
[
|
| [25] |
|
| [26] |
杨鑑初. 台湾温度初步分析[J]. 气象学报, 1950, 21(1): 47-52.
[
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
新疆植物志编辑委员会. 新疆植物志·简本[M]. 乌鲁木齐: 新疆人民出版总社, 新疆科技卫生出版社, 2014: 343-344.
[ Commissione Redactorum Florae Xinjianggensis. Flora Xinjiangensis (the Brief Edition)[M]. Urumqi: Xinjiang People’s Publishing House, Xinjiang Science and Technology Press, 2014: 343-344. ]
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}