
科尔沁沙地油松固沙林土壤真菌群落结构及功能
|
高海燕(1993-),女,主要从事荒漠化防治相关研究. E-mails: nmghy1993@163.com |
收稿日期: 2024-07-31
修回日期: 2024-09-29
网络出版日期: 2025-08-13
基金资助
内蒙古自治区“揭榜挂帅”项目(2024JBGS0005-4-2)
内蒙古自治区“揭榜挂帅”项目(2024JBGS0007)
Structure and function of soil fungal community in Pinus tabuliformis sand-fixing forests in Horqin Sandy Land
Received date: 2024-07-31
Revised date: 2024-09-29
Online published: 2025-08-13
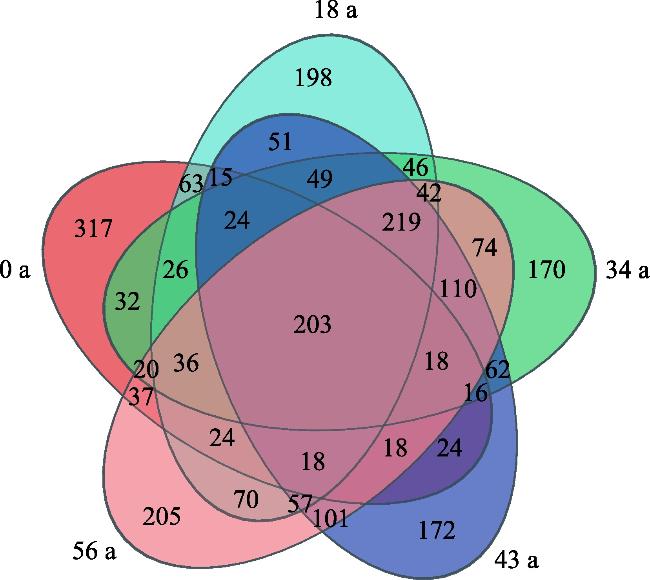
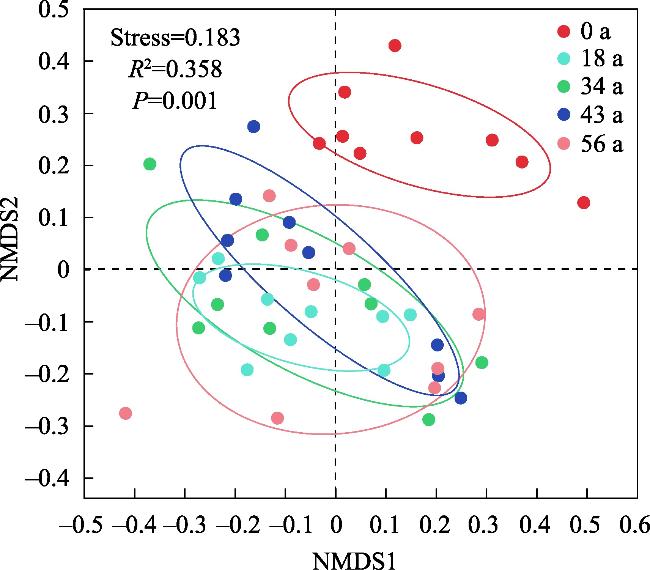
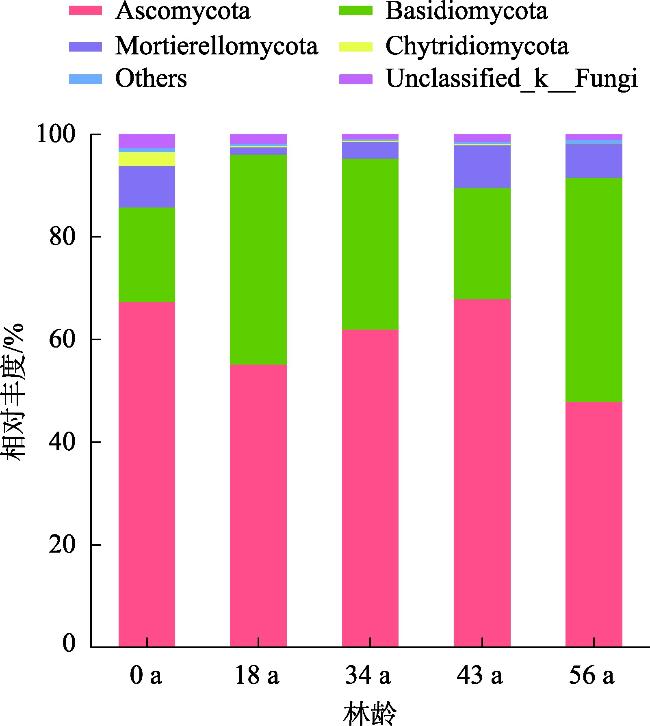
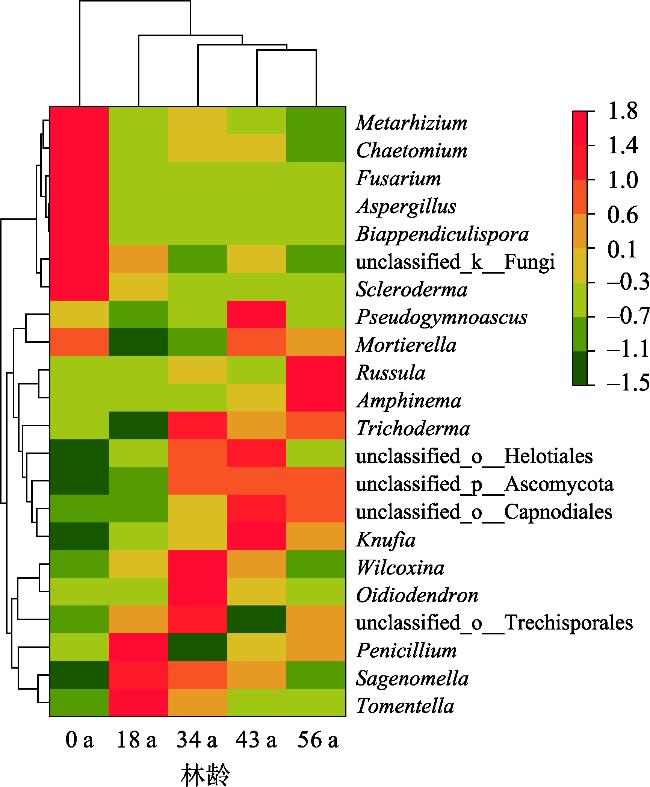
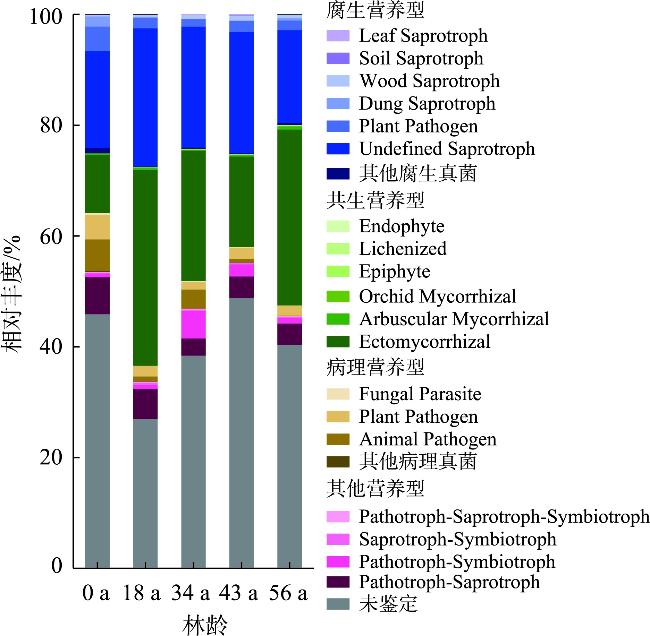
探究不同林龄油松固沙林土壤真菌群落结构及功能与土壤化学性质、酶活性的关系,为油松固沙林合理经营和管护提供理论依据。以科尔沁沙地流动沙地为对照(0 a),选择固沙18 a、34 a、48 a、56 a油松林地为研究对象,采用高通量测序技术解析土壤真菌群落结构和功能类群差异。结果表明:(1) 从油松固沙林土壤中获得2517个OTU,隶属于真菌的14个门、48个纲、127个目、286个科、579个属;优势菌门为子囊菌门(Ascomycota,47.91%~67.34%)、担子菌门(Basidiomycota,18.45%~43.70%)和被孢霉门(Mortierellomycota,1.41%~8.36%);优势属为Biappendiculispora、硬皮马勃属Scleroderma、棉革菌属Tomentella、Knufia、糙缘腺革菌属Amphinema。(2) 由韦恩图和NMDS分析表明,造林对土壤真菌群落结构影响较大,各林龄土壤真菌丰富度(ace指数、chao指数)显著增加(P<0.05),且与有机质、全氮、全磷、脲酶、脱氢酶、过氧化氢酶、中性磷酸酶、蔗糖酶、中性蛋白酶呈显著正相关关系(P<0.05)。(3) 真菌群落主要以共生营养型和腐生营养型为主,造林后共生营养型相对丰度均较对照增加,腐生营养型相对丰度较为稳定。造林对土壤真菌群落结构及功能发挥着重要的调节作用,研究结果丰富了固沙林土壤微生物群落研究内容,为科尔沁沙地油松固沙林土壤健康评价提供依据。
高海燕 , 张胜男 , 杨制国 , 张雷 , 黄海广 , 闫德仁 . 科尔沁沙地油松固沙林土壤真菌群落结构及功能[J]. 干旱区研究, 2025 , 42(1) : 118 -126 . DOI: 10.13866/j.azr.2025.01.11
The purpose of this study was to explore the relationship between the soil fungal community structure and function and soil chemical properties and enzyme activity in Pinus tabuliformis sand-fixing forests of different ages, so as to provide a theoretical basis for the rational management and protection of P. tabuliformis sand-fixing forests. Taking the mobile sand of Horqin Sandy Land as the control (0 year), P. tabuliformis forests with sand fixation for 18, 34, 48 and 56 years were selected as the research objects, and high-throughput sequencing technology was used to analyze the differences in soil fungal community structure and functional groups. The results show: (1) The 2517 OTUs obtained from the soil of the P. tabuliformis sand-fixing forest belong to 14 phyla, 48 classes, 127 orders, 286 families, and 579 genera of fungi. The dominant phyla were Ascomycota (47.91%-67.34%), Basidiomycota (18.45%-43.70%), and Mortierellomycota (1.41%-8.36%); the dominant genera were Biappendiculispora, Scleroderma, Tomentella, Knufia, and Amphinema. (2) Venn diagram and NMDS analysis showed that afforestation has a greater impact on soil fungal community structure. The ace index and chao index of soil fungi at each stand age increased significantly (P<0.05), and were related to organic matter, total nitrogen, and total phosphorus, urease, dehydrogenase, catalase, neutral phosphatase, sucrase and neutral protease have a significant positive correlation (P<0.05). (3) The fungal community was mainly composed of symbiotic and saprophytic types. After afforestation, the relative abundance of symbiotic types increased compared with the control, while the relative abundance of saprophytic types was relatively stable. Afforestation plays an important regulatory role on the structure and function of soil fungal communities. The research results enrich the research content of soil microbial communities in sand-fixing forests and provide a basis for soil health evaluation of P. tabuliformis sand-fixing forests in Horqin Sandy Land.
表1 不同处理土壤真菌群落Alpha多样性Tab. 1 Alpha diversity of soil fungal communities under different treatments |
| 林龄 | 覆盖度/% | Shannon指数 | Simpson指数 | Pielou指数 | chao指数 | ace指数 |
|---|---|---|---|---|---|---|
| 0 a | 99.95a | 2.918a | 0.147a | 0.547a | 247b | 244b |
| 18 a | 99.87b | 2.956a | 0.149a | 0.521a | 372a | 369a |
| 34 a | 99.87b | 2.958a | 0.127a | 0.537a | 360a | 365a |
| 43 a | 99.87b | 3.313a | 0.090a | 0.590a | 386a | 382a |
| 56 a | 99.85b | 2.936a | 0.211a | 0.504a | 436a | 426a |
注:同列不同小写字母表示不同处理间差异显著(P<0.05)。 |
表2 不同处理土壤养分特征Tab. 2 Soil nutrient characteristics of different treatments |
| 指标 | 林龄 | ||||
|---|---|---|---|---|---|
| 0 a | 18 a | 34 a | 43 a | 56 a | |
| 有机质/(g·kg-1) | 2.75±1.07d | 6.96±2.22c | 10.78±2.94b | 12.95±2.84a | 11.95±3.27ab |
| 全氮/(g·kg-1) | 0.23±0.06c | 0.43±0.11b | 0.56±0.19ab | 0.62±0.16a | 0.62±0.23a |
| 全磷/(g·kg-1) | 0.08±0.02b | 0.14±0.02a | 0.16±0.05a | 0.15±0.04a | 0.16±0.03a |
| 全钾/(g·kg-1) | 20.30±3.86a | 22.13±1.97a | 21.92±1.09a | 20.98±1.68a | 20.54±1.02a |
| 速效氮/(mg·kg-1) | 9.50±2.00ab | 6.28±2.91b | 12.61±2.87a | 12.19±1.96a | 11.29±2.45ab |
| 速效磷/(mg·kg-1) | 3.32±0.21b | 6.05±0.55a | 3.71±0.52b | 3.19±1.00b | 3.10±0.36b |
| 速效钾/(mg·kg-1) | 60.78±3.81b | 65.71±11.46b | 83.07±6.81a | 64.49±3.25b | 85.40±10.16a |
| 脲酶/(μg·g-1·h-1) | 5.39±0.50d | 7.82±0.33c | 8.15±0.89bc | 8.76±0.48b | 9.72±0.46a |
| 脱氢酶/(μg·g-1·h-1) | 0.07±0.00c | 0.25±0.09b | 0.27±0.09b | 0.33±0.05b | 0.52±0.19a |
| 过氧化氢酶/(mg·h-1·g-1) | 0.40±0.04b | 1.02±0.08a | 0.98±0.14a | 0.97±0.25a | 0.99±0.18a |
| 中性磷酸酶/(mg·h-1·g-1) | 56.76±7.28b | 111.92±44.63a | 108.21±24.31a | 111.76±14.17a | 126.57±25.14a |
| 蔗糖酶/(μg·h-1·g-1) | 94.42±4.17b | 180.67±37.13a | 136.75±26.26ab | 155.75±34.64ab | 187.22±57.95a |
| 中性蛋白酶/(μg·h-1·g-1) | 0.19±0.05c | 0.13±0.03c | 0.13±0.03c | 0.36±0.03b | 0.91±0.05a |
注:小写字母表示不同处理间差异显著(P<0.05)。 |
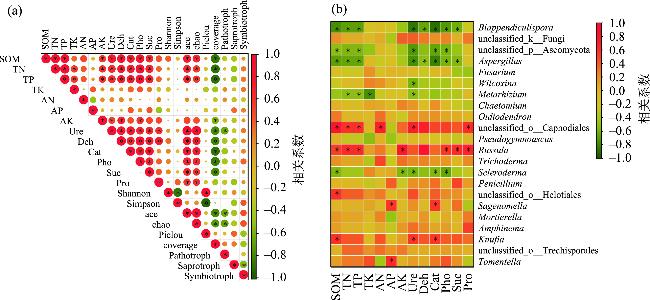
图6 土壤真菌多样性指数和功能类群(a)以及属水平真菌群落(b)与土壤养分的相关性分析注:*表示相关性显著,P<0.05;SOM为有机质,TN为全氮;TP为全磷;TK为全钾;AN为速效氮;AP为速效磷;AK为速效钾;Ure为脲酶;Deh为脱氢酶;Cat为过氧化氢酶;Pho为中性磷酸酶;Suc为蔗糖酶;Pro为中性蛋白酶。 Fig. 6 Correlation analysis between soil fungal diversity index and functional groups (a) and fungal communities at the genus level (b) and soil nutrients |
| [1] |
管鸿智, 黄荣珍, 王金平, 等. 红壤区退化林地表土真菌群落结构对土壤改良措施的响应[J]. 环境科学, 2023, 44(1): 494-501.
[
|
| [2] |
|
| [3] |
杨慧琴, 向涌旗, 吕倩, 等. 3种混交林造林初期土壤真菌群落结构特征[J]. 生态学报, 2024, 44(8): 3360-3371.
[
|
| [4] |
|
| [5] |
李丹丹, 李佳文, 高广磊, 等. 科尔沁沙地樟子松(Pinus sylvestris var. mongolia)人工林土壤真菌群落结构和功能特征[J]. 中国沙漠, 2023, 43(4): 241-251.
[
|
| [6] |
刘志民, 余海滨, 汪海洋. 科尔沁沙地“山水林田湖草沙”一体化治理原理及实施重点[J]. 应用生态学报, 2022, 33(12): 3441-3447.
[
|
| [7] |
高亮, 高永, 韩彦隆, 等. 不同治理措施下科尔沁沙地南缘土壤质地与持水特性研究[J]. 水土保持通报, 2018, 38(4): 1-6.
[
|
| [8] |
|
| [9] |
马义淑, 曹亚鑫, 牛敏, 等. 太岳山不同林龄人工油松林土壤微生物特征[J]. 环境科学, 2024, 45(4): 2406-2416.
[
|
| [10] |
逯军峰. 不同林龄油松人工林凋落物及其对土壤理化性质的影响研究[D]. 兰州: 甘肃农业大学, 2007.
[
|
| [11] |
|
| [12] |
刘亚栋, 王晓霞, 和璐璐, 等. 北京地区油松人工林不同演替类型空间结构对林下植被及土壤的影响[J]. 生态学报, 2023, 43(5): 1959-1970.
[
|
| [13] |
张从明, 满孝武, 吴芳, 等. 北京西山油松Pinus tabuliformis和樟子松P. varsylvestris. mongolica人工林土壤-林木连续体微生物群落比较[J]. 菌物学报, 2023, 42(10): 2041-2062.
[
|
| [14] |
张胜男, 高海燕, 闫德仁, 等. 科尔沁沙地油松人工林土壤细菌群落特征及影响作用[J]. 内蒙古林业科技, 2023, 49(4): 46-52.
[
|
| [15] |
章家恩. 生态学常用实验研究方法与技术[M]. 北京: 化学工业出版社, 2006.
[
|
| [16] |
徐林芳, 米媛婷, 柳兰洲, 等. 内蒙古不同类型草原土壤真菌群落结构及其影响因子的研究[J]. 草地学报, 2023, 31(7): 1977-1987.
[
|
| [17] |
王海英, 郭守玉, 黄满荣, 等. 子囊菌较担子菌具有更快的进化速率和更高的物种多样性[J]. 中国科学: 生命科学, 2010, 40(8): 731-737, 765-772.
[
|
| [18] |
马欣. 陇中半干旱区苜蓿种植年限对黄绵土微生物群落结构的影响解析[D]. 兰州: 甘肃农业大学, 2020.
[
|
| [19] |
|
| [20] |
王娅楠, 冯伟, 杨文斌. 低覆盖度行带式固沙林带间土壤微生物群落多样性[J]. 中国沙漠, 2024, 44(4): 115-123.
[
|
| [21] |
刘立玲, 周光益, 党鹏, 等. 湘西石漠化区3种造林模式土壤真菌群落结构差异[J]. 生态学报, 2022, 42(10): 4150-4159.
[
|
| [22] |
|
| [23] |
张健, 谷口武士, 徐明, 等. 盐碱培养条件对几种外生菌根真菌生长的影响[J]. 食用菌学报, 2021, 28(1): 61-65.
[
|
| [24] |
王华峰, 魏阳, 纪丽莲, 等. 不同绿僵菌定殖水稻根系及其促生效应[J]. 微生物学通报, 2024, 51(9): 3523-3534.
[
|
| [25] |
黄艳英, 彭晓辉, 欧桂宁, 等. 连作木薯对根际与非根际土壤真菌群落结构演替的影响[J]. 广西植物, 2024, 44(10): 1864-1877.
[
|
| [26] |
李小利. 外生菌根真菌菌株耐盐碱性筛选以及黑松(Pinus thunbergii Parl. )菌根苗合成耐盐碱性研究[D]. 南京: 南京农业大学, 2013.
[
|
| [27] |
|
| [28] |
|
| [29] |
曹鹏. 一株抗黄萎病的番茄内生放线菌的分离、筛选和鉴定[D]. 哈尔滨: 东北农业大学, 2016.
[
|
| [30] |
|
| [31] |
彭丽鸿, 崔朝伟, 王佳琪, 等. 连栽对杉木人工林土壤表层真菌群落结构及功能的影响[J]. 福建农林大学学报: 自然科学版, 2022, 51(5): 621-628.
[
|
| [32] |
|
| [33] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}