
模拟干湿条件下土壤酶活性对坝地土壤有机碳矿化的影响
|
邢欣然(2000-),男,硕士研究生,主要从事土壤侵蚀及碳循环研究. E-mail: 13099525526@163.com |
收稿日期: 2024-04-21
修回日期: 2024-06-12
网络出版日期: 2025-08-13
基金资助
宁夏自然科学基金项目(2023AAC03054)
国家自然科学基金项目(42473069)
Simulated effects of soil enzyme activity on soil organic carbon mineralization in dam land under dry and wet conditions
Received date: 2024-04-21
Revised date: 2024-06-12
Online published: 2025-08-13
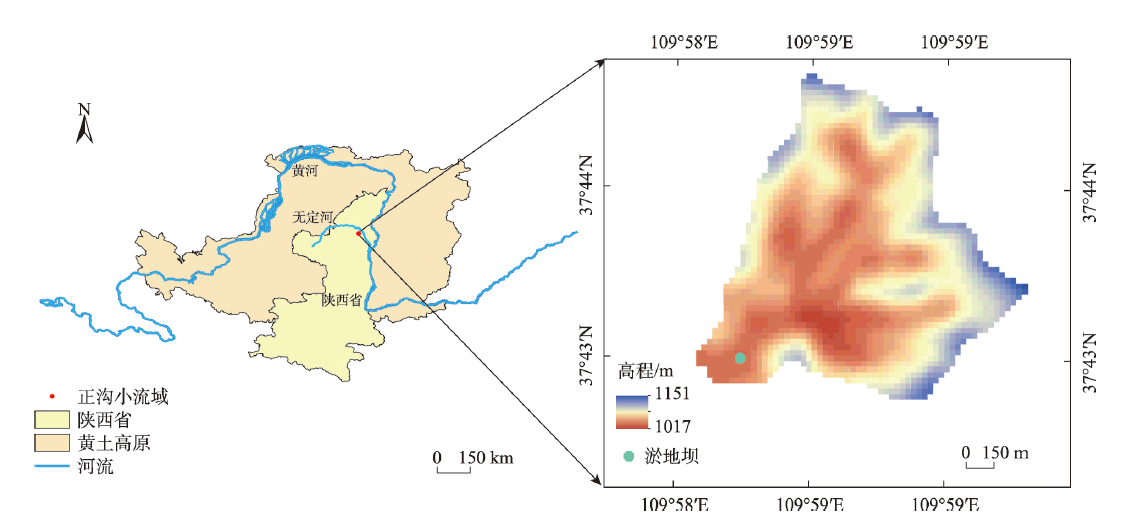
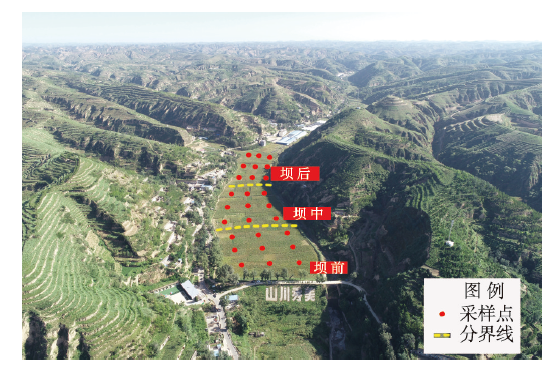
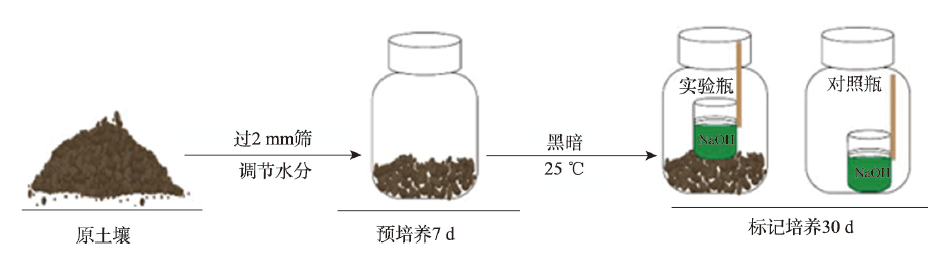
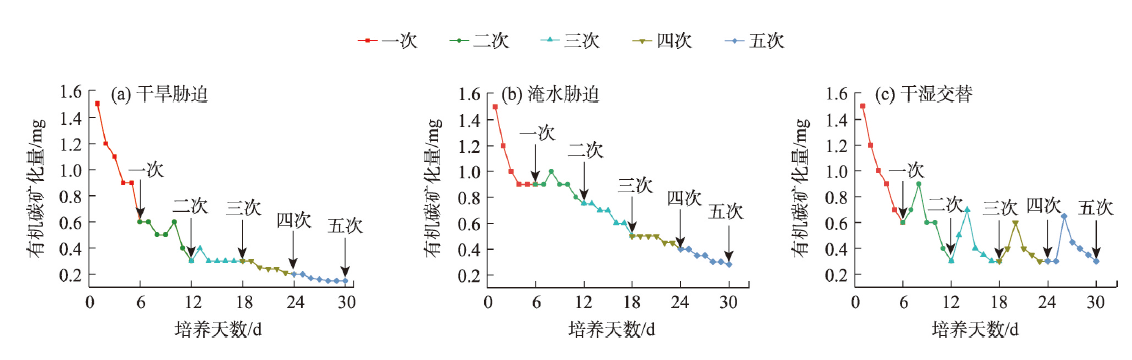
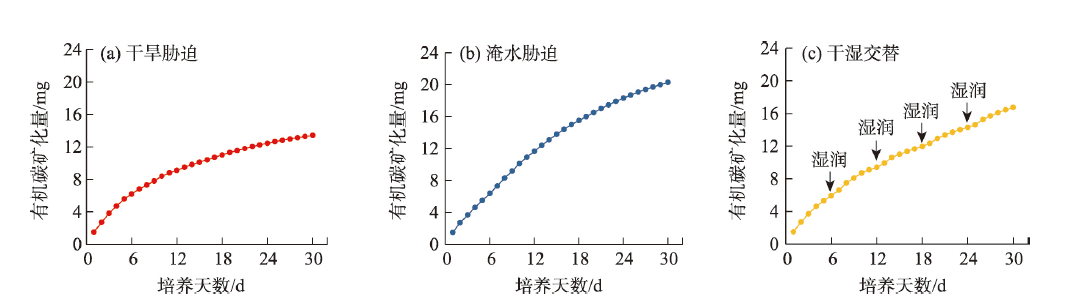
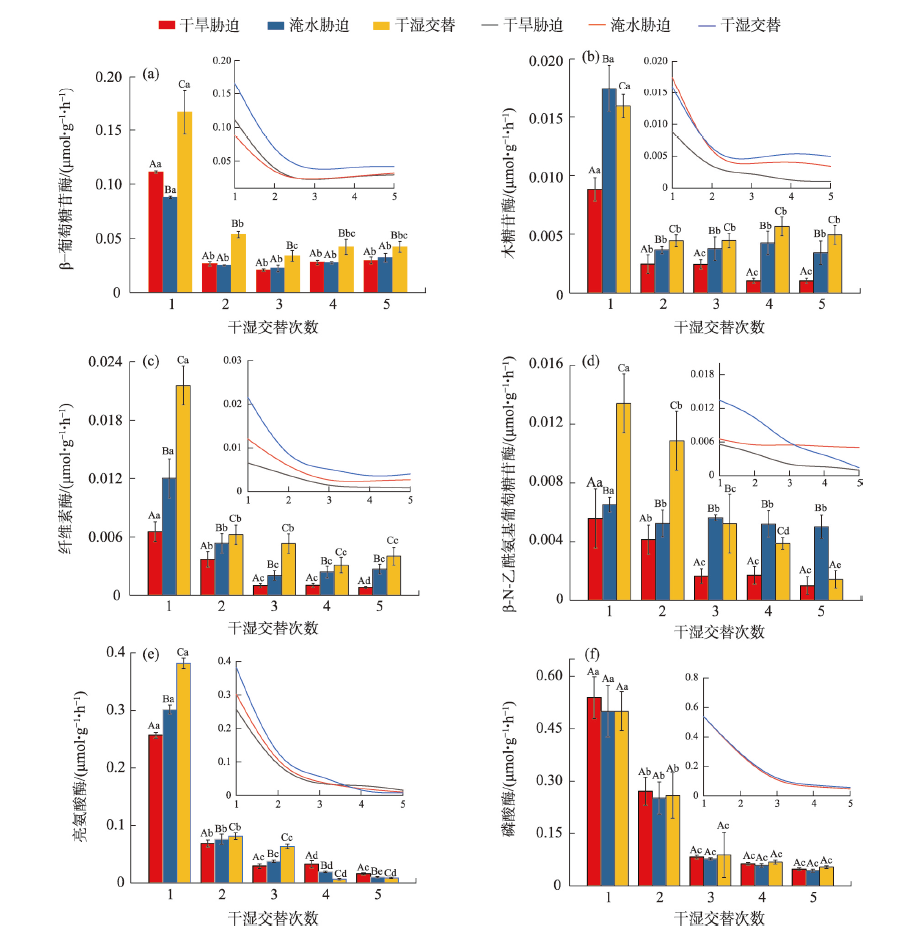
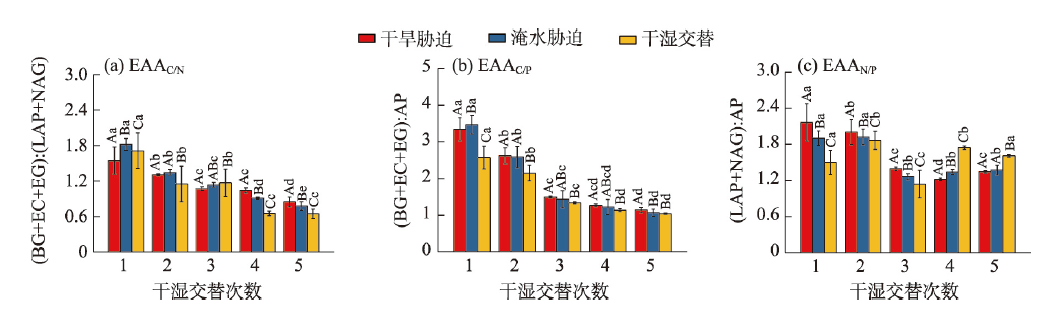
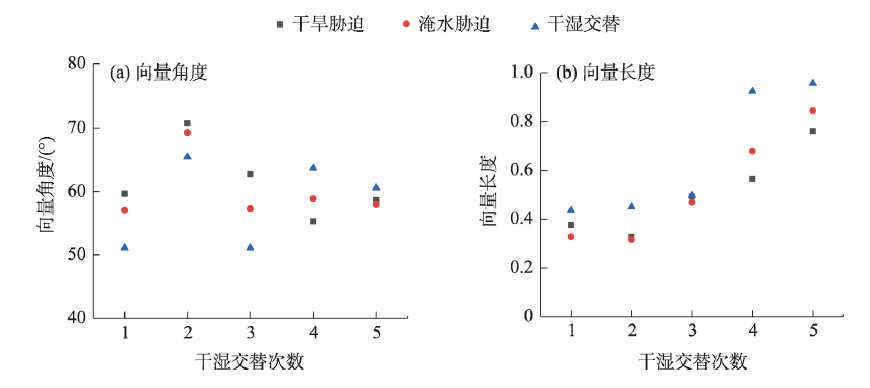
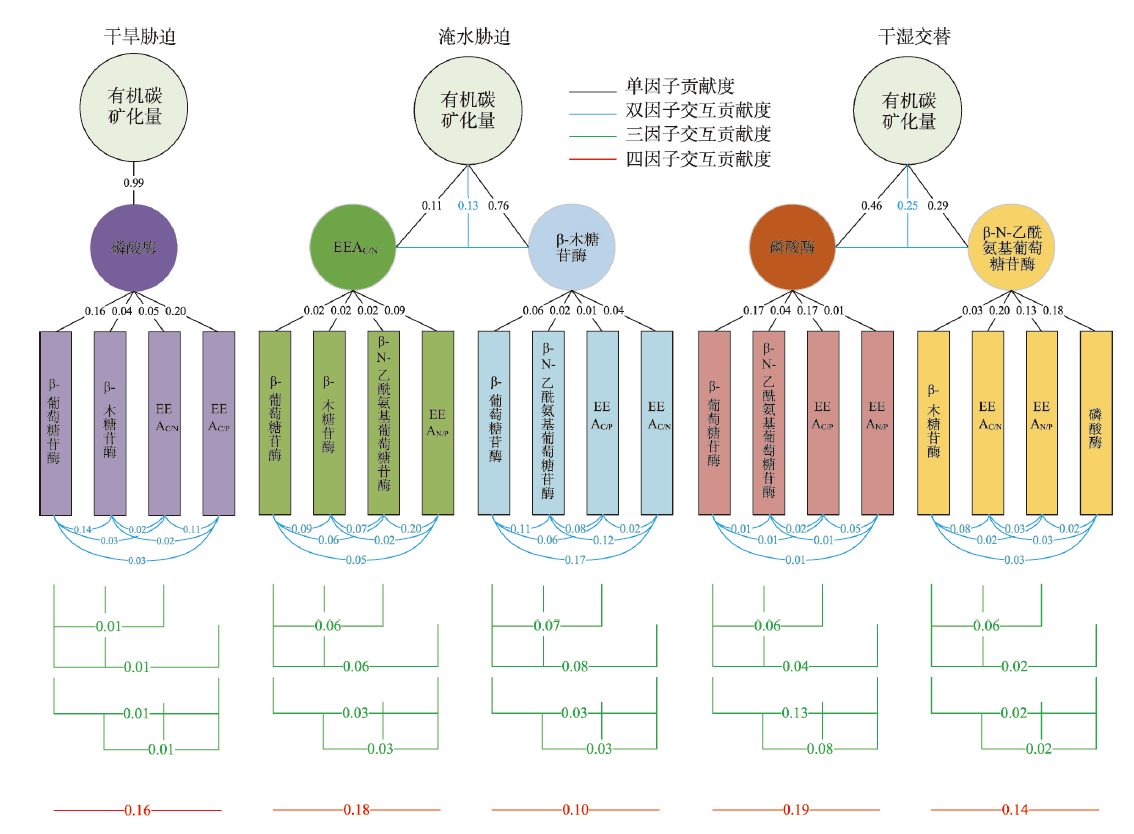
气候变化导致长期干旱和强降雨事件的发生频率增加,从而对生态系统碳循环产生影响。因此,了解不同干湿条件土壤酶活性如何影响有机碳矿化,能够帮助深入理解碳循环机制并推进全球碳中和目标达成。据此,本研究选择黄土高原特有的控制侵蚀所修建的淤地坝为研究对象,并设计淹水胁迫、干旱胁迫和干湿循环3种处理监测土壤酶活性及有机碳矿化过程。结果表明:干湿循环对有机碳矿化存在瞬时激发效应,且有机碳累积矿化量介于干旱胁迫和淹水胁迫之间。同时干湿循环均会增加碳循环和氮循环相关酶活性,但随着循环次数的增加酶活性逐渐降低并趋于稳定。三种水分处理条件下酶活性均受到碳和磷的限制,而干湿循环次数越多碳限制越强烈,第四次干湿交替后干湿交替条件下磷限制作用超过淹水胁迫和干旱胁迫。当土壤处于干旱、淹水胁迫以及干湿循环时,限制有机碳矿化的酶活性因子也各不相同。干旱胁迫下磷酸酶因子的直接作用为99%,淹水胁迫下EAAC/N(碳循环相关酶/氮循环相关酶)和木糖苷酶因子的直接作用为87%,干旱胁迫和淹水胁迫下因子间交互作用仅为1%和13%。干湿循环条件下,磷酸酶和β-N-乙酰氨基葡萄糖苷酶因子的直接作用为75%,因子间交互作用为25%,干湿循环可以显著增加因子之间的相互联系。本研究可为明晰黄土高原淤地坝建设在国家碳中和目标中发挥的作用提供理论支撑。
邢欣然 , 张祎 , 李鹏 , 刘晓君 , 陶清瑞 , 任正龑 , 胥世斌 . 模拟干湿条件下土壤酶活性对坝地土壤有机碳矿化的影响[J]. 干旱区研究, 2024 , 41(11) : 1969 -1980 . DOI: 10.13866/j.azr.2024.11.16
Climate change has increased the frequency of long-term droughts and heavy rainfall events, impacting ecosystems’ carbon cycle. Therefore, understanding how soil enzyme activity in different dry/wet conditions affects organic carbon mineralization can help deepen our understanding of the carbon cycle mechanism and advance the goal of global carbon neutrality. This study was based on the unique terraces built to control erosion in the Loess Plateau, and three treatments of flooding stress, drought stress, and wet/dry cycles were designed to monitor soil enzyme activity and organic carbon mineralization. The results indicate that the wet/dry cycle has a transient stimulating effect on organic carbon mineralization, and the cumulative mineralized organic carbon occurs between the drought and flooding stress. At the same time, the wet/dry cycle will increase the activity of carbon and nitrogen cycle-related enzymes; however, the enzyme activity will gradually decrease and stabilize as the number of cycles increases. The enzyme activity in all three water treatment conditions was limited by carbon and phosphorus, and the carbon limitation was more substantial with increasing wet/dry cycles. After the fourth alternating wet/dry cycle, the phosphorus limitation exceeded the flooding and drought stress treatments. When the soil was under drought stress, flooding stress, or wet/dry cycles, the enzyme activity factors that limit organic carbon mineralization were different. The direct effect of the phosphatase factor under drought stress was 99%, the direct impact of EAAC/N (carbon cycle- /nitrogen cycle-related enzyme) and xyloglucosidase factor under flooding stress was 87%, and the direct effect of the interaction between factors under drought and flooding stress was only 1% and 13%. Under wet/dry cycle conditions, the direct impact of phosphatase and N-acetyl-beta-glucosaminidase factor was 75%, the direct effect of interaction between factors was 25%, and the wet/dry cycle significantly increased the interaction between factors. This study provides theoretical support for clarifying the role of Loess Terrace.
表1 土壤理化性质Tab. 1 Physicochemical properties of soil |
| 类型 | 粉粒 /% | 砂粒 /% | 黏粒 /% | 土壤含 水率/% | 有机碳 /(g·kg-1) | 全氮 /(g·kg-1) | 全磷 /(g·kg-1) | 速效磷 /(mg·kg-1) | 铵态氮 /(mg·kg-1) | 硝态氮 /(mg·kg-1) |
|---|---|---|---|---|---|---|---|---|---|---|
| 坝地 | 74.52±7.84 | 14.93±7.96 | 10.53±0.15 | 34.50±3.89 | 4.52±0.93 | 1.11±0.34 | 3.01±0.33 | 0.018±0.010 | 1.574±0.140 | 0.012±0.010 |
注:数据为平均值±标准差。 |
表2 土壤酶活性种类、缩写、底物和功能Tab. 2 Types of soil enzyme activities their abbreviations, substrate and functions |
| 土壤酶 | 缩写 | 底物 | 类型 | 功能 |
|---|---|---|---|---|
| β-葡萄糖苷酶 | BG | 4-MUB-β-D-glucoside | C-targeting h ydrolytic | 碳循环酶 |
| 木糖苷酶 | EC | 4-Methylumbelliferone-β-D-xylopyranoside | C-targeting hydrolytic | 碳循环酶 |
| 纤维素水解酶 | EG | 4-Methylumbelliferyl-Beta-D-cellobiopyranoside | C-targeting hydrolytic | 碳循环酶 |
| β-N-乙酰氨基葡萄糖苷酶 | NAG | β-N-Acetylglucosaminidase | N-targeting hydrolytic | 氮循环酶 |
| 亮氨酸酶 | LAP | L-Leucine-7-amido-4-methylcoumarin hydrochloride | N-targeting hydrolytic | 氮循环酶 |
| 磷酸酶 | AP | 4-Methylumbelliferyl phosphate | P-targeting hydrolytic | 磷循环酶 |
表3 有机碳矿化量与酶活性含量的逐步回归分析Tab. 3 Stepwise regression analysis of organic carbon mineralization and enzyme activity levels |
| 类型 | 逐步回归模型 | 标准系数 | R2 | |
|---|---|---|---|---|
| 预测变量 | 模型方程 | |||
| 干旱胁迫 | 磷酸酶 | Y=1.636X+0.118 | 0.989 | 0.972 |
| 淹水胁迫 | EAAC/N | Y=0.923X1-16.291X2-0.323 | 1.143 | 0.997 |
| β-木糖苷酶 | -0.174 | |||
| 干湿交替 | 磷酸酶 | Y=1.682X1-20.951X2+0.351 | 1.385 | 0.998 |
| β-N-乙酰氨基葡萄糖苷酶 | -0.421 | |||
| [1] |
周波涛. 全球气候变暖: 浅谈从AR5到AR6的认知进展[J]. 大气科学学报, 2021, 44(5): 667-671.
[
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
刘峰, 赵鹏程, 张昀, 等. 微生物角度揭示气候变暖对土壤有机碳转化影响的研究综述[J]. 土壤通报, 2022, 53(6): 1492-1498.
[
|
| [7] |
|
| [8] |
张睿媛, 袁丹, 秦树平, 等. 碳氮磷化学计量比对土壤有机碳矿化激发效应的影响[J]. 中国生态农业学报(中英文), 2023, 31(8): 1311-1321.
[
|
| [9] |
赵宇航, 殷浩凯, 胡雪纯, 等. 长期秸秆还田褐土有机碳矿化特征及其驱动力[J]. 环境科学, 2024, 45(4): 2353-2362.
[
|
| [10] |
于淑华, 张丽霞, 谢雪迎, 等. 同水分模式对山东茶园土壤氮素动态的影响[J]. 土保持学报, 2021, 35(4): 289-298.
[
|
| [11] |
杨媛媛, 李占斌, 高海东, 等. 大理河流域淤地坝拦沙贡献率分析[J]. 水土保持学报, 2021, 35(1): 85-89.
[
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
王君, 宋新山, 王苑. 多重干湿交替对土壤有机碳矿化的影响[J]. 环境科学与技术, 2013, 36(11): 31-35.
[
|
| [21] |
高俊琴, 徐兴良, 张锋, 等. 水分梯度对若尔盖高寒湿地土壤活性有机碳分布的影响[J]. 水土保持学报, 2008, 22(3): 126-131.
[
|
| [22] |
陈玉军, 李婷, 朱立安, 等. 湛江红树林湿地不同淹水梯度下土壤养分及其化学计量特征[J]. 西北林学院学报, 2023, 38(5): 19-27.
[
|
| [23] |
张红星, 王效科, 冯宗炜, 等. 黄土高原小麦田土壤呼吸对强降雨的响应[J]. 生态学报, 2008, 28(12): 6189-6196.
[
|
| [24] |
|
| [25] |
|
| [26] |
马伟伟, 王丽霞, 李娜, 等. 不同水氮水平对川西亚高山林地土壤酶活性的影响[J]. 生态学报, 2019, 39(19): 7218-7228.
[
|
| [27] |
肖如武, 黄楚龙, 宗钊辉, 等. 低磷胁迫对烤烟根系有机酸含量及土壤磷酸酶活性的影响[J]. 广东农业科学, 2021, 48(8): 74-82.
[
|
| [28] |
王梅, 晏梓然, 赵子文, 等. 黄土高原植被演替过程中相对土壤酶活性的变化特征[J]. 水土保持学报, 2021, 35(5): 181-187.
[
|
| [29] |
|
| [30] |
潘新雅, 李军保, 陈阳, 等. 6个紫花苜蓿品种根系形态结构对低磷胁迫的响应[J]. 草地学报, 2021, 29(11): 2494-2504.
[
|
| [31] |
|
| [32] |
阮长明, 唐国勇, 杜寿康, 等. 金沙江干热河谷不同海拔土壤碳氮磷化学计量和酶活性研究[J]. 西南农业学报, 2023, 36(11): 2464-2472.
[
|
| [33] |
刘亚军, 吴娟, 邹锋, 等. 鄱阳湖湿地灰化薹草洲滩土壤微生物和酶特性对水分梯度的响应[J]. 湿地科学, 2017, 15(2): 269-275.
[
|
| [34] |
宋霄君, 吴会军, 武雪萍, 等. 长期保护性耕作可提高表层土壤碳氮含量和根际土壤酶活性[J]. 植物营养与肥料学报, 2018, 24(6): 1588-1597.
[
|
| [35] |
梅孔灿, 陈岳民, 范跃新, 等. 凋落叶和磷添加对马尾松林土壤碳激发效应的影响[J]. 土壤学报, 2022, 59(4): 1089-1099.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}