
新疆野苹果叶绿体基因组特征及其进化历史
|
张健(1999-),男,硕士研究生,主要从事干旱区植物保护遗传学研究. E-mail: zj13201654032@163.com |
收稿日期: 2024-01-04
修回日期: 2024-10-06
网络出版日期: 2025-08-12
基金资助
国家自然科学基金(32170391)
中国科学院战略生物资源计划(KFJ-BRP-007-008)
中国科学院“西部青年学者”项目(2022-XBQNXZ-007)
Characteristics and evolutionary history of the chloroplast genome in Malus sieversii
Received date: 2024-01-04
Revised date: 2024-10-06
Online published: 2025-08-12
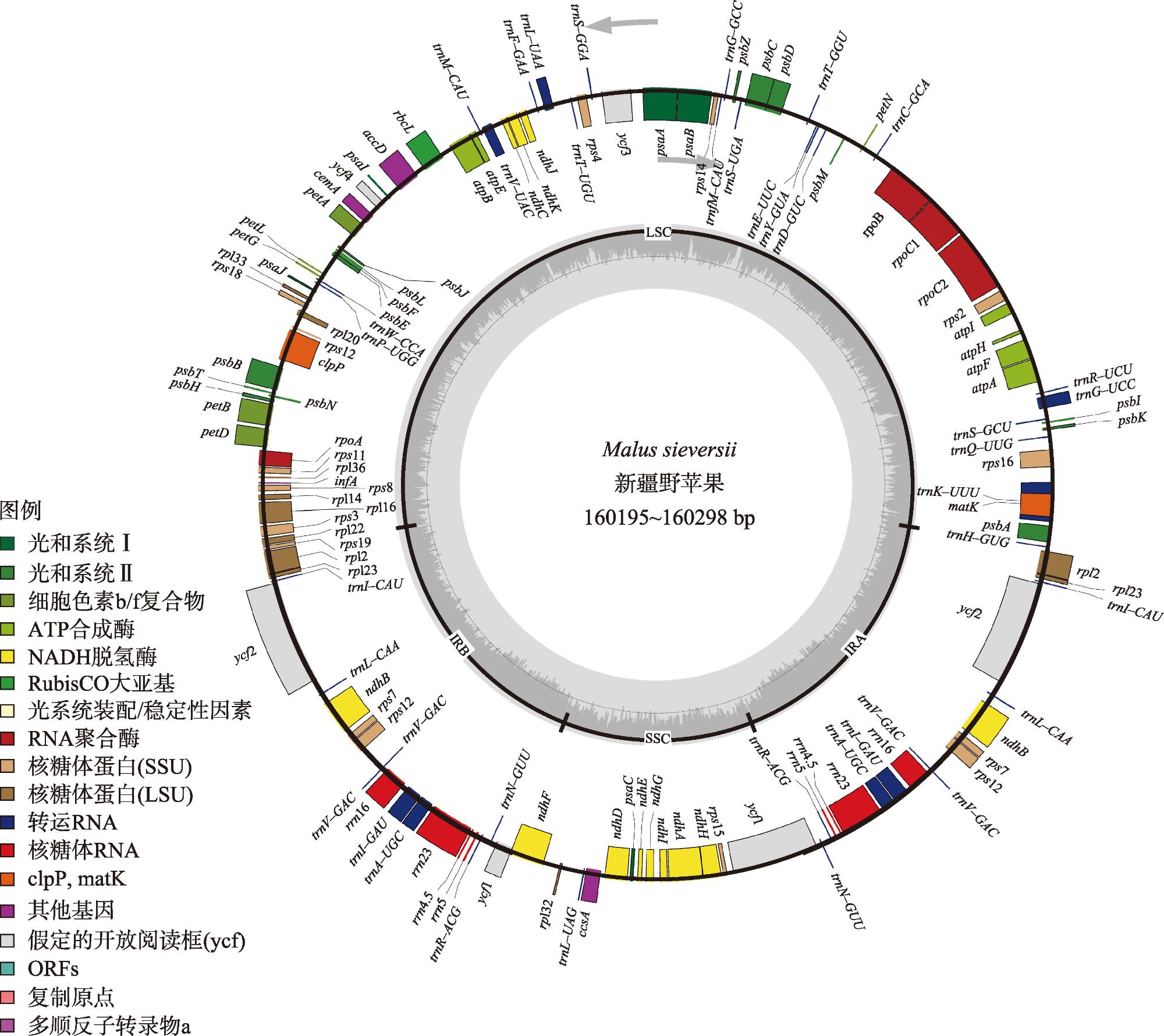
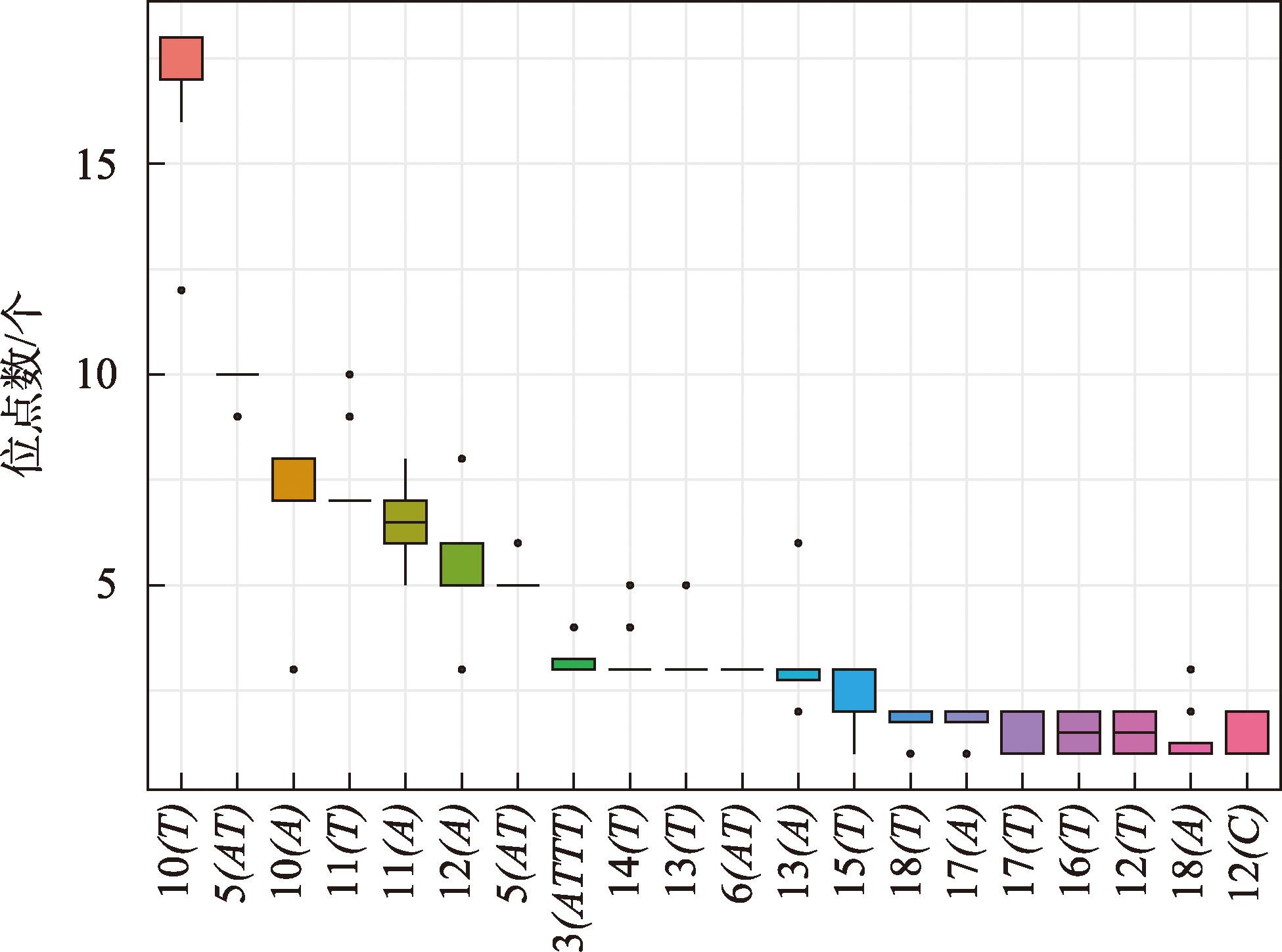
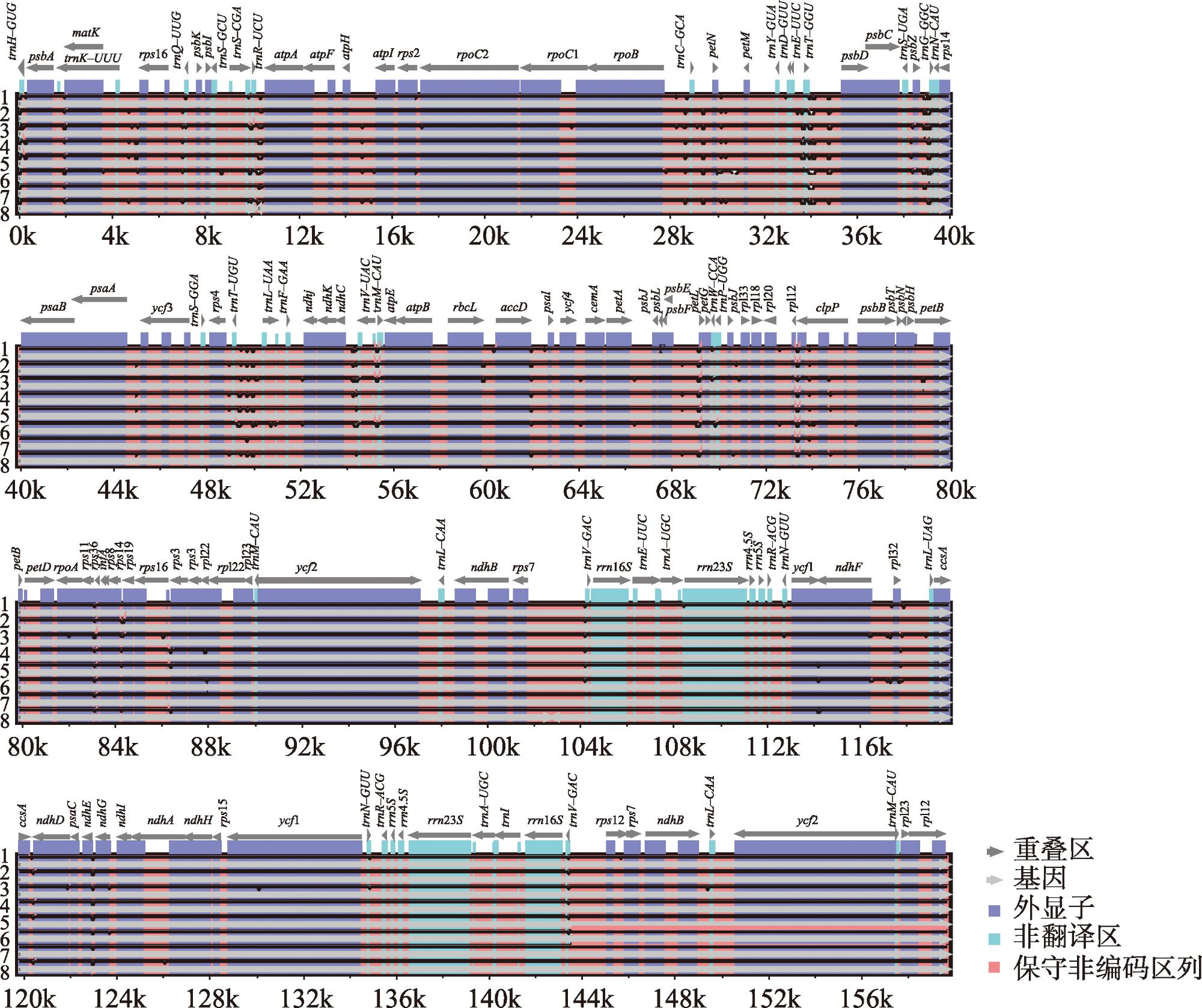
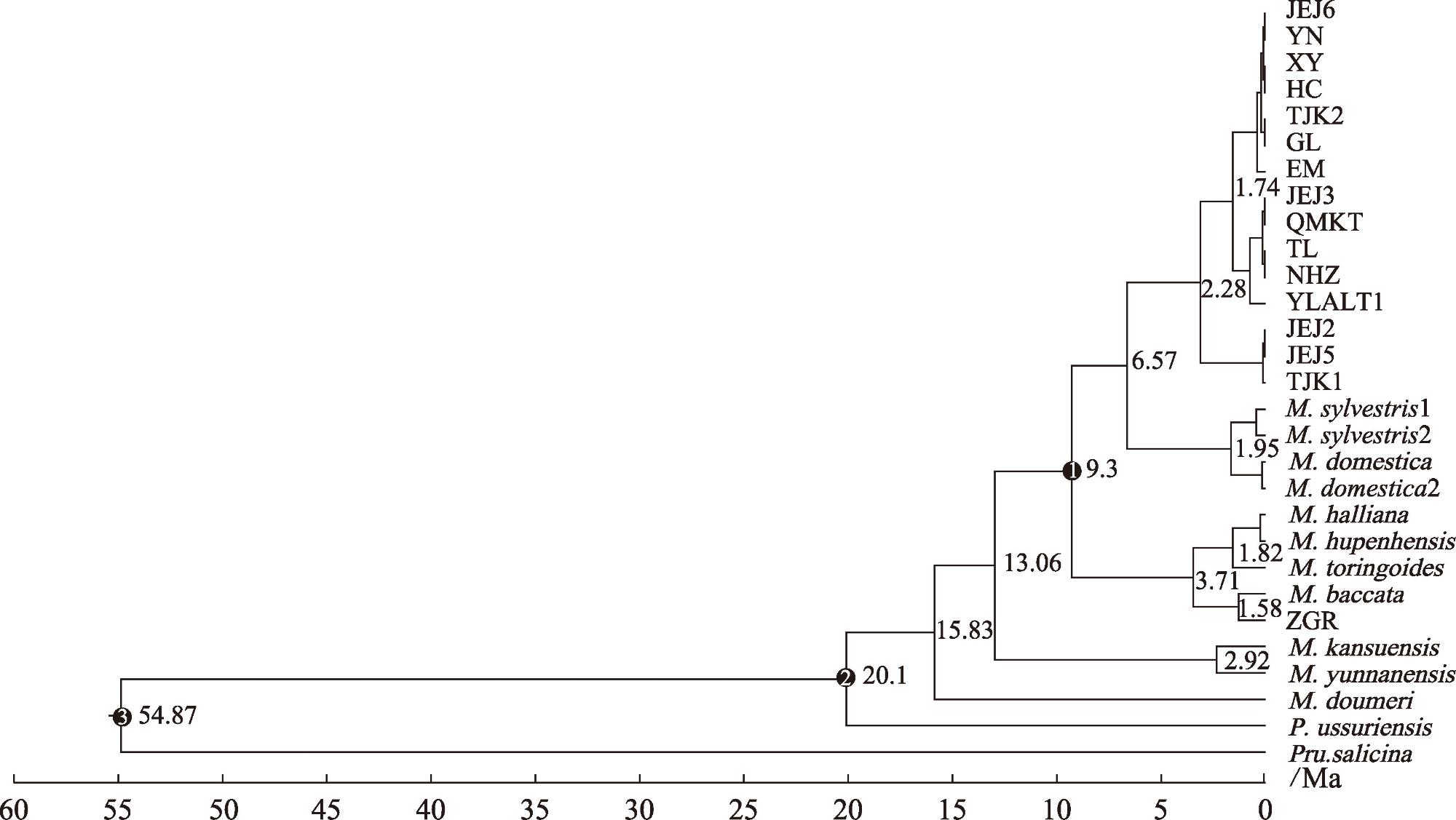
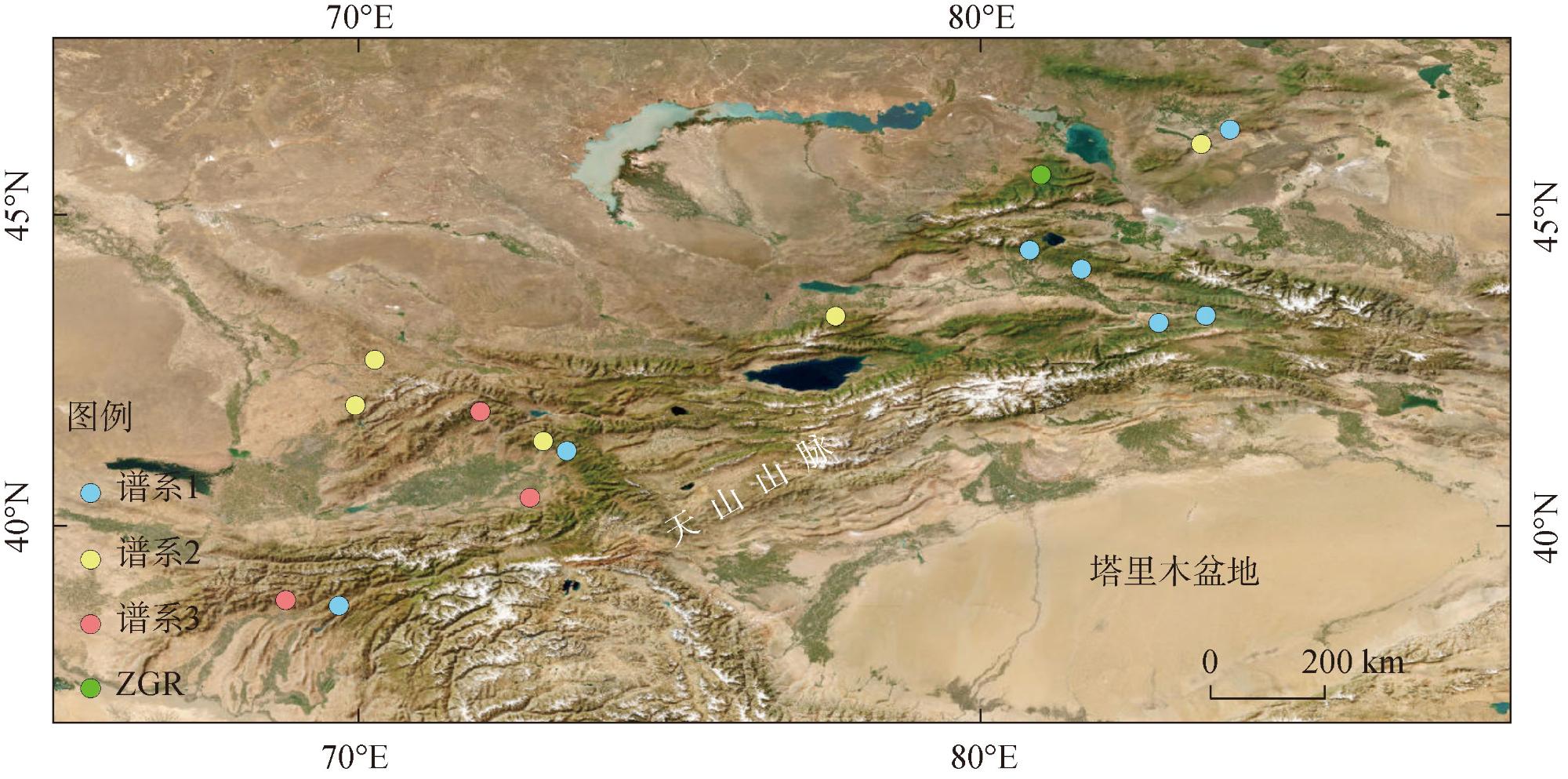
新疆野苹果是苹果属重要的种质资源,属国家二级保护植物,同时也是栽培苹果的祖先之一。通过比较新疆野苹果不同种群的叶绿体基因组结构特征差异,来阐明新疆野苹果谱系分化格局与物种演化历史。利用Illumina NovaSeq平台对来自16个不同地区的种群进行了全基因组测序,每个种群选取了一个代表性个体。测序数据经过质量控制后,进行了基因组组装和功能注释。随后,对组装得到的基因组进行了深入的结构分析和谱系分化研究。结果显示:新疆野苹果叶绿体基因组序列全长160195~160279 bp,具有典型的四分结构。叶绿体基因组共注释到131个基因;检测出长重复序列48~58个,简单重复序列93~101个。新疆野苹果与苹果属其他物种的叶绿体基因组在IR(反向重复)区域的变异程度较低,而检测到的变异主要发生在非编码区域。系统发育方面,新疆野苹果最终分成了三个谱系,谱系Ⅰ主要分布在东部,谱系Ⅱ与谱系Ⅲ主要分布在西部。谱系Ⅰ与谱系Ⅱ的分化时间为1.74 Ma,谱系Ⅰ、谱系Ⅱ与谱系Ⅲ的分化时间为2.28 Ma。新疆野苹果遗传分化受到了第四纪气候变化的影响。相较于国外分布的新疆野苹果,中国分布的新疆野苹果遗传多样性较低,对于分布在中国的新疆野苹果需分别采取不同的方式保护,并着重关注遗传多样性较高的塔城区域。
张健 , 张宏祥 . 新疆野苹果叶绿体基因组特征及其进化历史[J]. 干旱区研究, 2024 , 41(12) : 2110 -2119 . DOI: 10.13866/j.azr.2024.12.12
Malus sieversii, a state-protected species and the progenitor of cultivated apples, is an important germplasm resource within the genus Malus. In this study, we aimed to compare the structural characteristics of chloroplast genomes across various populations of M. sieversii, clarify the lineage divergence pattern, and trace the evolutionary history of this species. We used the Illumina NovaSeq platform to conduct whole-genome sequencing of individuals from 16 different populations, with one sample representing each population. After conducting quality control on the sequencing data, we conducted genome assembly and functional annotation. Subsequently, we conducted a comprehensive structural analysis and lineage differentiation studies on the assembled genomes. The chloroplast genome length in M. sieversii ranged from 160195 to 160279 base pairs (bp), exhibiting a typical tetrad structure. In total, 131 genes were identified within the chloroplast genome, along with 48-58 long repeats and 93-101 simple sequence repeats. Notably, variations in the IR region between M. sieversii and other species in the genus were minimal, predominantly occurring in noncoding regions. Phylogenetic analysis revealed that M. sieversii clusters into three distinct lineages: lineage I, primarily occupying the eastern part of the distribution range, and lineages II and III, predominantly found in the west. The divergence time between lineages I and II was approximately 1.74 million years ago (Ma), while the divergence between lineages I, II, and III was around 2.28 Ma. These findings indicate that the lineage divergences of M. sieversii were significantly influenced by climate changes during the Quaternary period. Compared to internationally distributed populations, M. sieversii in China shows relatively low genetic diversity. Therefore, tailored conservation strategies should be implemented for M. sieversii across different regions, with particular emphasis on protecting genetically diverse populations in the Tacheng area.
表1 新疆野苹果样品采集信息Tab. 1 Sample collection information of Malus sieversii |
| 种群编号 | 国家来源 | 经度 | 纬度 |
|---|---|---|---|
| EM | 中国 | 84.00°E | 46.36°N |
| GL | 中国 | 82.86°E | 43.26°N |
| HC | 中国 | 80.78°E | 44.43°N |
| JEJ2 | 吉尔吉斯斯坦 | 71.95°E | 41.84°N |
| JEJ3 | 吉尔吉斯斯坦 | 72.96°E | 41.36°N |
| JEJ5 | 吉尔吉斯斯坦 | 72.75°E | 40.45°N |
| JEJ6 | 吉尔吉斯斯坦 | 73.34°E | 41.21°N |
| NHZ | 哈萨克斯坦 | 70.26°E | 42.67°N |
| QMKT | 哈萨克斯坦 | 69.95°E | 41.94°N |
| TJK1 | 塔吉克斯坦 | 68.82°E | 38.81°N |
| TJK2 | 塔吉克斯坦 | 69.68°E | 38.72°N |
| TL | 中国 | 83.54°E | 46.13°N |
| XY | 中国 | 83.61°E | 43.38°N |
| YLALT | 哈萨克斯坦 | 77.67°E | 43.37°N |
| YN | 中国 | 81.62°E | 44.12°N |
| ZGR | 哈萨克斯坦 | 80.96°E | 45.64°N |
表2 新疆野苹果叶绿体基因组中含内含子的基因Tab. 2 Genes with introns in the chloroplast genome of Malus sieversii |
| 基因名称 | 基因数 量/个 | 内含子 数量/个 | 基因组 |
|---|---|---|---|
| ndhA | 1 | 1 | NADH脱氢酶的亚基 |
| ndhB | 2 | 1 | NADH脱氢酶的亚基 |
| petB | 1 | 1 | 细胞色素b/f复合体的亚基 |
| petD | 1 | 1 | 细胞色素b/f复合体的亚基 |
| atpF | 1 | 1 | ATP合酶的亚基 |
| rpl16 | 1 | 1 | 大核糖体亚基的蛋白质 |
| rpl2 | 2 | 1 | 大核糖体亚基的蛋白质 |
| rps12 | 2 | 2 | 小核糖体亚基的蛋白质 |
| rps16 | 1 | 1 | 小核糖体亚基的蛋白质 |
| rpoC1 | 1 | 1 | RNA聚合酶的亚基 |
| trnA-UGC | 2 | 1 | 转运RNA |
| trnG-GCC | 1 | 1 | 转运RNA |
| trnG-UCC | 1 | 1 | 转运RNA |
| trnK-UUU | 1 | 1 | 转运RNA |
| trnL-UAA | 1 | 1 | 转运RNA |
| trnT-UGU | 1 | 1 | 转运RNA |
| trnV-UAC | 1 | 1 | 转运RNA |
| trnS-GCU | 1 | 1 | 转运RNA |
| clpP | 1 | 2 | 蛋白酶 |
| ycf3 | 1 | 2 | 保守的假设性叶绿体开放阅读框 |
| [1] |
|
| [2] |
张新时. 伊犁野果林的生态地理特征和群落学问题[J]. 植物学报, 1973, 15(2): 239-253.
[
|
| [3] |
|
| [4] |
米尔卡米力·麦麦提, 刘忠权, 马晓东, 等. 新疆野苹果的生存现状、问题及保护策略[J]. 广西植物, 2021, 41(12): 2100-2109.
[
|
| [5] |
张宏祥, 郑田勇. 生境片段化对新疆野苹果种群遗传结构的影响[J]. 干旱区研究, 2020, 37(3): 715-721.
[
|
| [6] |
|
| [7] |
|
| [8] |
赵玉芬. 叶绿体基因组在植物学研究中的应用进展[J]. 生物学教学, 2022, 47(3): 83-85.
[
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
Geneious Prime[EB/OL]. https://www.geneious.com
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
崔大方, 廖文波, 羊海军, 等. 中国伊犁天山野果林区系表征地理成分及区系发生的研究[J]. 林业科学研究, 2006, 19(5): 555-560.
[
|
| [29] |
刘兴诗, 林培钧, 钟骏平. 伊犁野果林生境分析和发生探讨[J]. 干旱区研究, 1993, 10(3): 28-33.
[
|
| [30] |
张宏祥, 闻志彬, 王茜. 新疆野苹果种群遗传结构及其环境适应性[J]. 植物生态学报, 2022, 46(9): 1098-1108.
[
|
| [31] |
周小东, 常顺利, 王冠正, 等. 天山北坡中段雪岭云杉径向生长对气候变化的响应[J]. 干旱区研究, 2023, 40(8): 1215-1228.
[
|
| [32] |
赵卓怡, 郝兴明. 基于Priestley-Taylor方法的中亚干旱区实际蒸散特征及归因[J]. 干旱区研究, 2023, 40(7): 1085-1093.
[
|
| [33] |
褚佳瑶, 冯琳骄, 侯毅兴, 等. 新疆野苹果种群受损现状[J]. 经济林研究, 2022, 40(1): 265-273.
[
|
| [34] |
董翰林, 王文婷, 谢云, 等. 新疆气候干湿变化特征及其影响因素[J]. 干旱区研究, 2023, 40(12): 1875-1884.
[
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}