
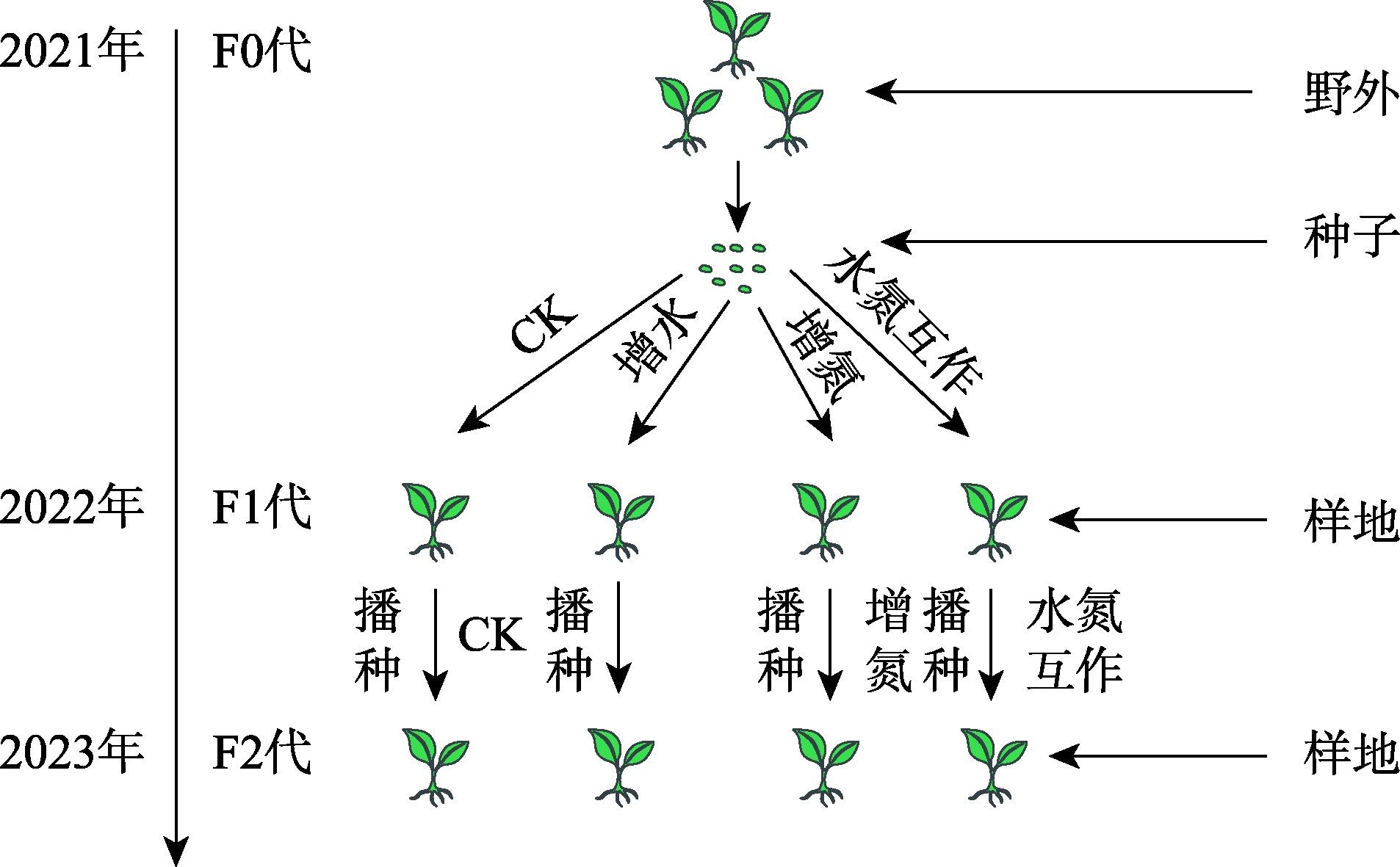
降水、氮沉降对尖喙牻牛儿苗隔代生理可塑性的影响
|
沙涛(1996-),男,硕士研究生,主要从事荒漠植物生态适应研究. E-mail: 1796379306@qq.com |
收稿日期: 2024-04-21
修回日期: 2024-05-15
网络出版日期: 2025-08-12
基金资助
国家自然科学基金项目(32160256)
国家自然科学基金项目(32171513)
国家自然科学基金项目(31971428)
新疆维吾尔自治区天山英才项目(2023TSYCCX0082)
Effects of precipitation and nitrogen deposition on transgenerational plasticity in alternate generations of Erodium oxyrhinchum
Received date: 2024-04-21
Revised date: 2024-05-15
Online published: 2025-08-12
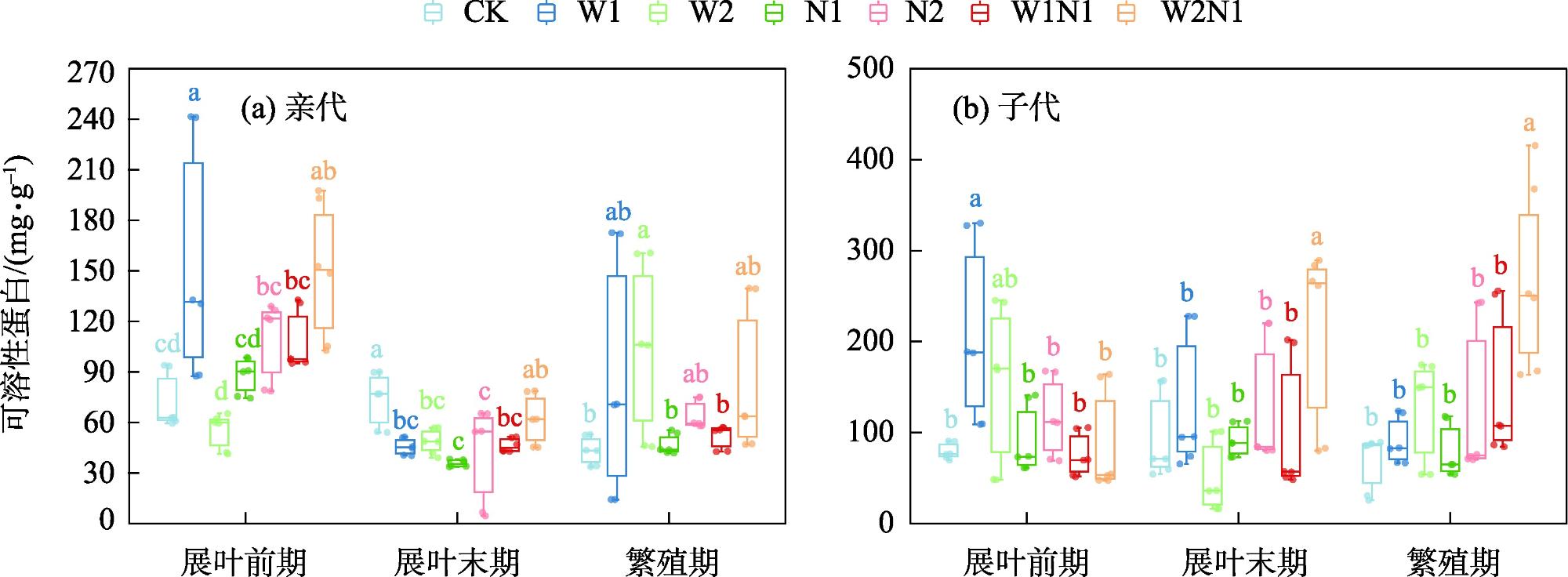
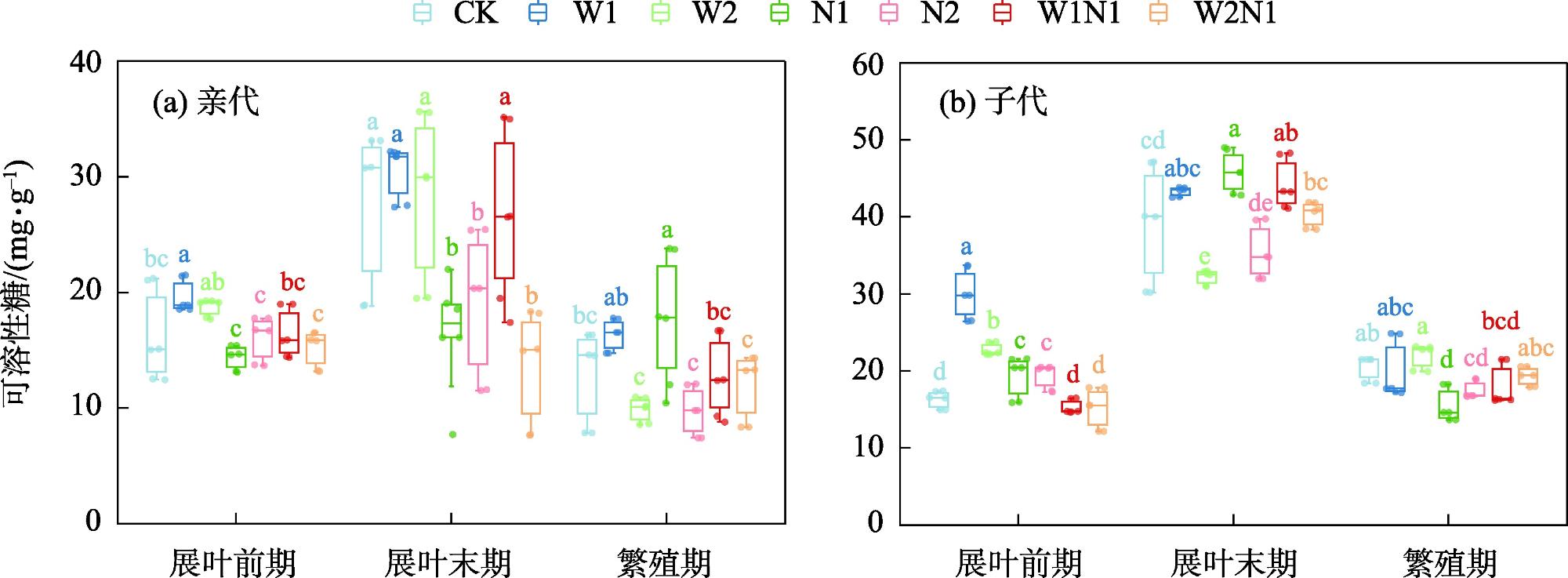
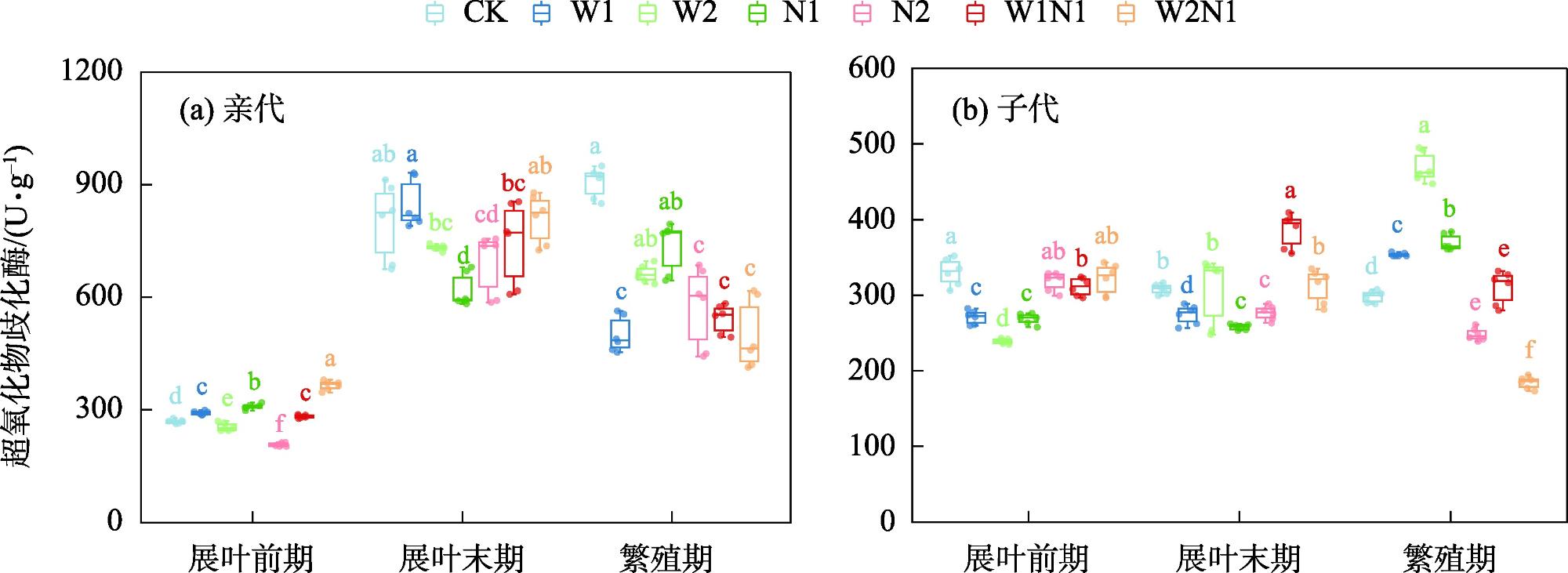
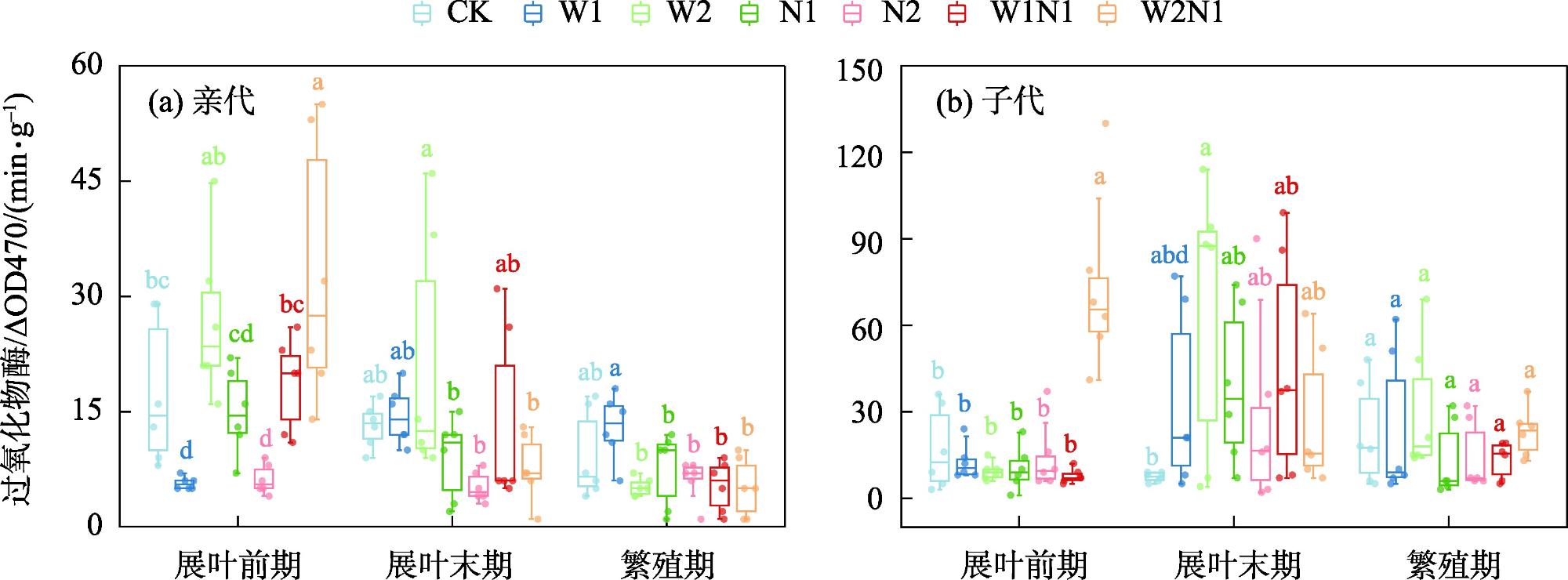
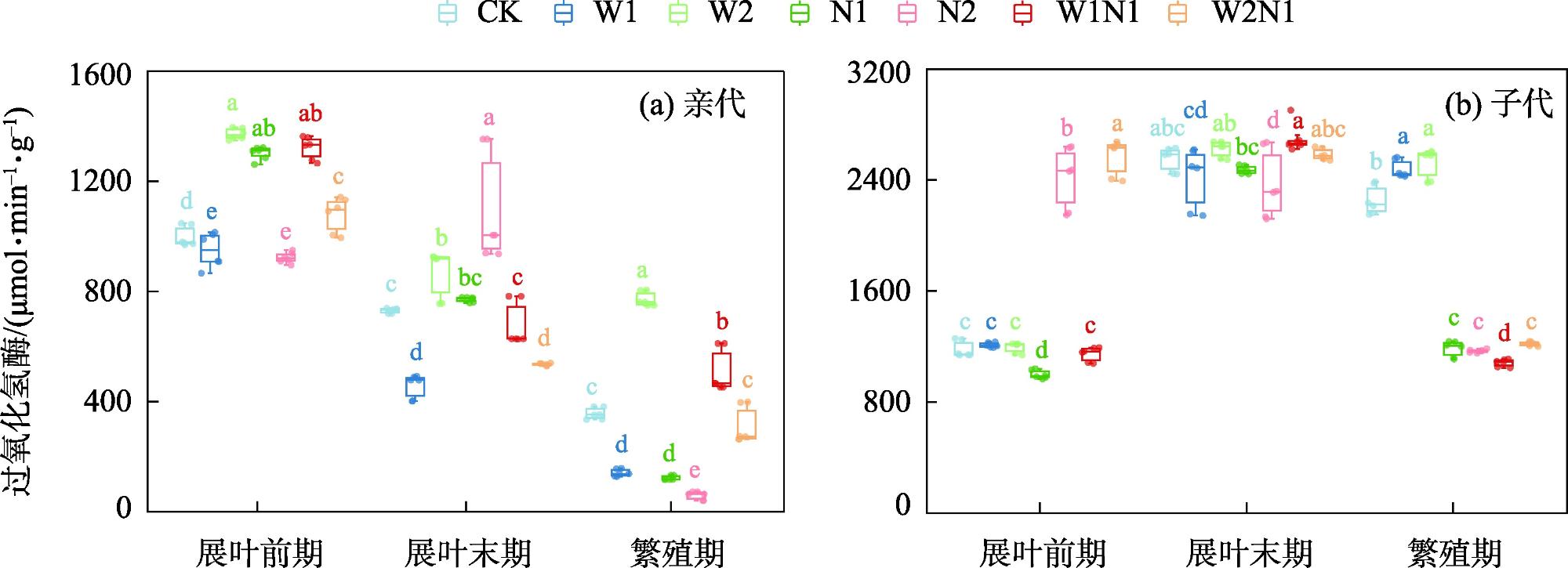
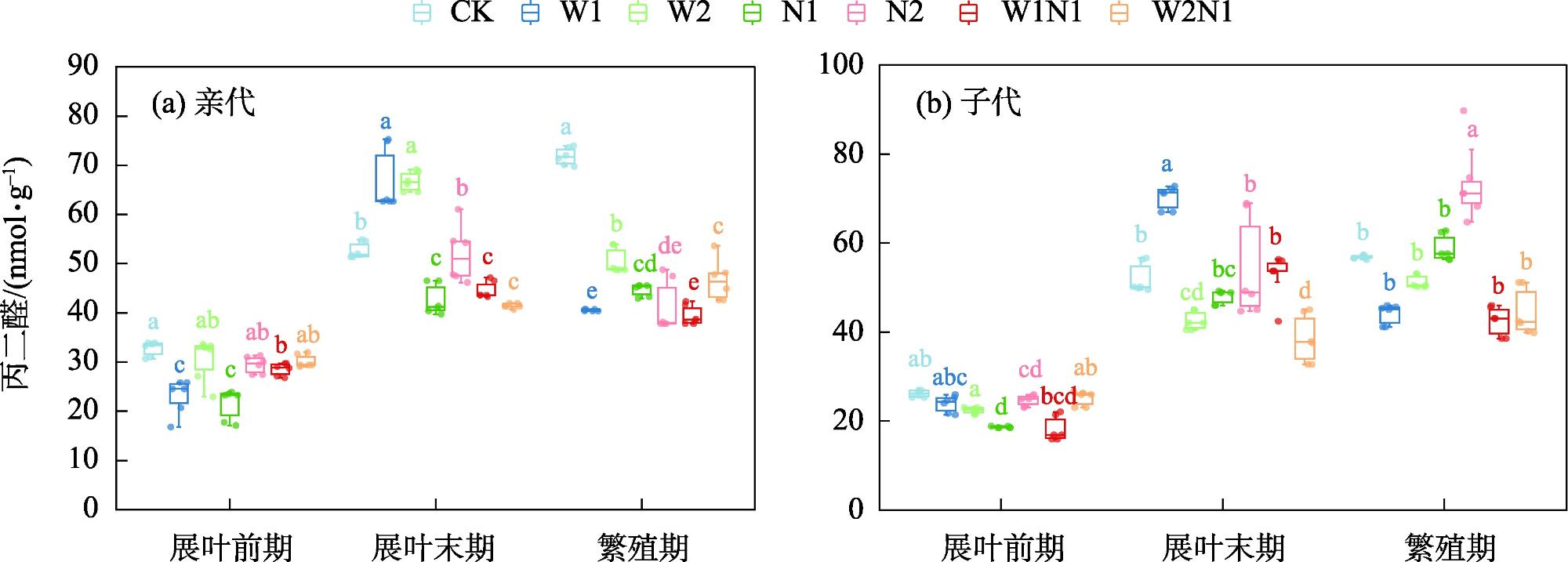
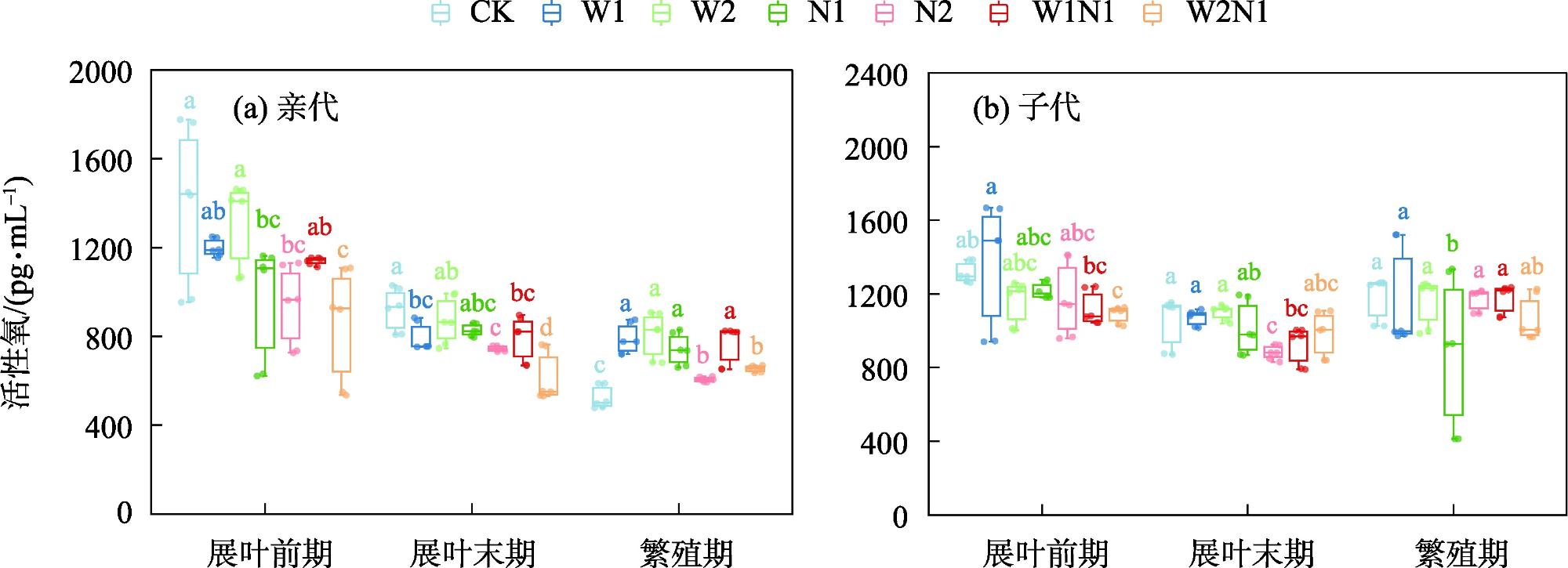
隔代可塑性是后代免受环境胁迫的有效方式,通过增加子代与环境匹配的可能性来缓冲环境变化对子代的影响,提高子代对所处环境的适应度。因此,本研究以生长发育快、对环境响应敏感的一年生短命植物优势种尖喙牻牛儿苗(Erodium oxyrhinchum)作为研究材料,结合增水、增氮处理,从短命植物生理生化指标角度分析短命植物代际间对气候因子的响应差异,尝试明确气候因子对荒漠短命植物隔代生理可塑性的影响。结果表明:(1) 增水处理显著提高了两代植株的可溶性糖(SS)含量、其余生理指标呈代际差异性,增水处理下可溶性蛋白(SP)、可溶性糖(SS)、过氧化物酶(POD)呈隔代适应性。(2) 增氮处理显著抑制亲代植株的超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、丙二醛(MDA)含量,对子代植株过氧化氢酶(CAT)、活性氧(ROS)含量呈抑制作用,增氮处理下可溶性蛋白(SP)、超氧化物歧化酶(SOD)、过氧化物酶(POD)呈隔代适应性。(3) 水氮互作处理显著促进亲代植株的过氧化物酶(POD)、过氧化氢酶(CAT)、活性氧(ROS)含量,但显著抑制超氧化物歧化酶(SOD)含量。对子代植株超氧化物歧化酶(SOD)、过氧化物酶(POD)含量呈促进作用,过氧化氢酶(CAT)、丙二醛(MDA)、活性氧(ROS)含量呈抑制作用,水氮互作处理下可溶性蛋白(SP)、过氧化物酶(POD)呈隔代适应性。总体而言,降水、氮沉降对尖喙牻牛儿苗隔代生理可塑性具有显著影响,尖喙牻牛儿苗生理生化指标对增水、增氮以及水氮互作处理呈现不同的响应差异。未来降水、氮沉降的增加可能改变尖喙牻牛儿苗后代的生态适应能力和策略,进而改变其种群未来发展趋势。
沙涛 , 张玲卫 , 刘会良 , 张岚 , 卢妤婷 , 周鑫宇 , 文晓虎 , 张元明 . 降水、氮沉降对尖喙牻牛儿苗隔代生理可塑性的影响[J]. 干旱区研究, 2024 , 41(10) : 1753 -1766 . DOI: 10.13866/j.azr.2024.10.13
Transgenerational plasticity is an effective method to protect offspring from environmental stress, buffering the impact of environmental changes on the offspring by increasing the probability of matching the offspring with the environment and improving the adaptability of the offspring to the environment in which they live. Therefore, this study was conducted to analyze the transgenerational differences in the response of ephemeral plants to climatic factors from the perspective of their physiological and biochemical indexes and to elucidate the effects of climatic factors on the physiological plasticity of desert ephemeral plants in transgenerational periods. For this purpose, we used Erodium oxyrhinchum, a dominant species of annual ephemeral plants with rapid growth and development and sensitive response to the environment, in combination with water and nitrogen increase treatments. We also clarified the effect of climatic factors on the physiological plasticity of desert ephemeral plants across generations. Results showed that (1) the water addition treatment significantly increased the soluble sugar (SS) content of two generations of plants, the remaining physiological indexes exhibited transgenerational differences, and the soluble protein (SP), SS, and peroxidase (POD) exhibited transgenerational adaptations under the water addition treatment. (2) The nitrogen enrichment treatment significantly inhibited the production of superoxide dismutase (SOD), POD, catalase (CAT), and malondialdehyde (MDA) in the parental plants and inhibited the production of CAT and reactive oxygen species (ROS) in the offspring plants, and the contents of SP, SOD, and POD exhibited transgenerational adaptations. (3) The water-nitrogen interaction treatment significantly promoted the production of POD, CAT, and ROS in the progeny plants but significantly suppressed the production of SOD. The production of SOD and POD was promoted, and that of CAT, MDA, and ROS was inhibited in offspring plants. SP and POD were spaced-adapted under the water-nitrogen interaction treatment. Overall, precipitation and nitrogen deposition exerted a significant effect on the transgenerational physiological plasticity of E. oxyrhinchum, and the physiological and biochemical indexes of E. oxyrhinchum exhibited different response differences to the treatment of water increase, nitrogen increase, and water-nitrogen interactions. Increased precipitation and nitrogen deposition in the future may alter the ecological adaptive capacity and strategy of the descendants of E. oxyrhinchum, which may in turn alter the future development trend of their populations.
表1 增水、增氮及其互作对尖喙牻牛儿苗渗透调节物质响应值的影响Tab. 1 Effects of water addition, nitrogen addition and their interactions on response values of osmoregulatory substances in Erodium oxyrhinchum |
| 处理 | 可溶性蛋白SP | 可溶性糖SS | |||
|---|---|---|---|---|---|
| F1 | F2 | F1 | F2 | ||
| W1 | 0.3189 | 0.5283 | 0.1749 | 0.2285 | |
| W2 | 0.0638 | 0.2227 | -0.0328 | 0.0698 | |
| N1 | -0.1502 | 0.0866 | -0.1066 | 0.0171 | |
| N2 | 0.0802 | 0.4445 | -0.2197 | -0.0267 | |
| W1N1 | 0.0393 | 0.2733 | -0.0126 | -0.0224 | |
| W2N1 | 0.4036 | 0.7607 | -0.2817 | -0.0324 | |
注:W1为增水30%;W2为增水50%;N1为中N;N2为高N;W1N1为增水30%+中N;W2N1为增水50%+中N。下同。 |
表2 增水、增氮及其互作对尖喙牻牛儿苗抗氧化酶响应值的影响Tab. 2 Effect of water addition, nitrogen addition and their interactions on antioxidant enzyme response values of Erodium oxyrhinchum |
| 处理 | 超氧化物歧化酶SOD | 过氧化物酶POD | 过氧化氢酶CAT | |||||
|---|---|---|---|---|---|---|---|---|
| F1 | F2 | F1 | F2 | F1 | F2 | |||
| W1 | -0.1526 | -0.0488 | -0.2241 | 0.4255 | -0.4819 | 0.0223 | ||
| W2 | -0.1535 | 0.0421 | 0.1154 | 0.6329 | 0.4200 | 0.0502 | ||
| N1 | -0.1108 | -0.0573 | -0.2346 | 0.2150 | -0.2493 | -0.2810 | ||
| N2 | -0.2868 | -0.1104 | -0.7884 | 0.2260 | -0.4863 | -0.0045 | ||
| W1N1 | -0.1792 | 0.0679 | -0.1516 | 0.1735 | 0.1882 | -0.2362 | ||
| W2N1 | -0.0953 | -0.1649 | -0.1584 | 0.9334 | -0.1227 | 0.0585 | ||
表3 增水、增氮及其互作对尖喙牻牛儿苗氧化应激指标响应值的影响Tab. 3 Effects of water addition, nitrogen addition and their interactions on response values of oxidative stress indicators in Erodium oxyrhinchum |
| 处理 | 丙二醛MDA | 活性氧ROS | |||
|---|---|---|---|---|---|
| F1 | F2 | F1 | F2 | ||
| W1 | -0.2280 | -0.0163 | 0.0384 | 0.0148 | |
| W2 | -0.0626 | -0.1543 | 0.1055 | -0.0333 | |
| N1 | -0.3693 | -0.1297 | -0.0423 | -0.1320 | |
| N2 | -0.2245 | 0.1457 | -0.1523 | -0.0992 | |
| W1N1 | -0.3005 | -0.2153 | 0.0104 | -0.0985 | |
| W2N1 | -0.2511 | -0.1968 | -0.2215 | -0.1193 | |
| [1] |
|
| [2] |
陈娟, 张小晶, 李巧玉, 等. 不同坡向川西亚高山林木竞争与叶片表型可塑性的关系研究[J]. 生态学报, 2022, 42(5): 1788-1797.
[
|
| [3] |
王丹, 李江飞, 李亚麒, 等. 不同苗龄云南松异速生长及其表型可塑性[J]. 中南林业科技大学学报, 2022, 42(1): 36-44.
[
|
| [4] |
王玉萍, 高会会, 张峰, 等. 珠芽蓼叶片对海拔变化的表型可塑性[J]. 应用生态学报, 2021, 32(6): 2070-2078.
[
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
冯淑华, 陈雅君, 周阳. 干旱胁迫下三叶草渗透调节物质动态及与叶片水分的相关性[J]. 中国草地学报, 2011, 33(6): 70-74.
[
|
| [11] |
刘志芳, 张春梅, 闫治斌, 等. 干旱胁迫与复水对玉米苗期渗透调节物质及抗氧化酶的影响[J]. 甘肃农业科技, 2014, 45(1): 3-6.
[
|
| [12] |
杨舒贻, 陈晓阳, 惠文凯, 等. 逆境胁迫下植物抗氧化酶系统响应研究进展[J]. 福建农林大学学报(自然科学版), 2016, 45(5): 481-489.
[
|
| [13] |
孙岩, 何明珠, 王立. 降水控制对荒漠植物群落物种多样性和生物量的影响[J]. 生态学报, 2018, 38(7): 2425-2433.
[
|
| [14] |
闫建成, 梁存柱, 付晓玥, 等. 草原与荒漠一年生植物性状对降水变化的响应[J]. 草业学报, 2013, 22(1): 68-76.
[
|
| [15] |
赵娟, 宋媛, 孙涛, 等. 红松和蒙古栎种子萌发及幼苗生长对升温与降水综合作用的响应[J]. 生态学报, 2012, 32(24): 7791-7800.
[
|
| [16] |
|
| [17] |
毛祖美, 张佃民. 新疆北部早春短命植物区系纲要[J]. 干旱区研究, 1994, 11(3): 1-26.
[
|
| [18] |
张立运, 陈昌笃. 论古尔班通古特沙漠植物多样性的一般特点[J]. 生态学报, 2002, 22(11): 1923-1932.
[
|
| [19] |
陈艳锋. 模拟降水和氮沉降变化对短命植物生活史的影响[D]. 北京: 中国科学院大学, 2019.
[
|
| [20] |
卡吾沙尔·库都斯, 刘会良, 张岚, 等. 尖喙牻牛儿苗春/秋萌植株及子代种子的生理生化特性[J]. 干旱区研究, 2022, 39(5): 1473-1485.
[
|
| [21] |
黄韵杰, 李永刚, 尹本丰, 等. 齿肋赤藓(Syntrichia caninervis)氮磷计量特征对降水量的响应[J]. 中国沙漠, 2023, 43(2): 1-10.
[
|
| [22] |
段呈, 吴玲, 王绍明, 等. 近30年古尔班通古特沙漠短命植物的时空格局[J]. 生态学报, 2017, 37(8): 2642-2652.
[
|
| [23] |
|
| [24] |
周晓兵, 张元明. 干旱半干旱区氮沉降生态效应研究进展[J]. 生态学报, 2009, 29(7): 3835-3845.
[
|
| [25] |
覃磊, 郭磊, 张小波, 等. ROS信号对植物抗性的调控作用研究进展[J]. 激光生物学报, 2018, 27(3): 193-198.
[
|
| [26] |
张清航, 张永涛. 植物体内丙二醛(MDA)含量对干旱的响应[J]. 林业勘察设计, 2019, 48(1): 110-112.
[
|
| [27] |
林选栋, 武文莉, 林丽果, 等. 不同盐胁迫水平下硅对高羊茅幼苗生物量、酶活性和渗透调节物质的影响[J]. 草业科学, 2018, 35(7): 1653-1660.
[
|
| [28] |
韩冰, 贺超兴, 郭世荣, 等. 丛枝菌根真菌对盐胁迫下黄瓜幼苗渗透调节物质含量和抗氧化酶活性的影响[J]. 西北植物学报, 2011, 31(12): 2492-2497.
[
|
| [29] |
曲波, 陈鲜, 张婷, 等. 入侵植物瘤突苍耳(Xanthium strumarium)种子萌发期表型代间传递可塑性研究[J]. 沈阳农业大学学报, 2019, 50(6): 664-671.
[
|
| [30] |
师雅鑫, 王泽, 任财, 等. 不同施氮水平下梭梭幼苗对干旱胁迫的生理响应研究[J]. 广东农业科学, 2024, 51(1): 41-50.
[
|
| [31] |
周萍, 陈志国, 庄丽, 等. 水氮互作对滴灌春小麦渗透调节物及产量的影响[J]. 石河子大学学报(自然科学版), 2013, 31(4): 425-429.
[
|
| [32] |
褚妍, 任菲, 赵贵林, 等. 渗透胁迫对植物抗氧化酶影响的研究进展[J]. 安徽农业科学, 2011, 39(3): 1300-1302.
[
|
| [33] |
林宇丰, 李魏, 戴良英. 抗氧化酶在植物抗旱过程中的功能研究进展[J]. 作物研究, 2015, 29(3): 326-330.
[
|
| [34] |
吴玲, 李志辉, 吴际友, 等. 干旱胁迫对青冈栎种源叶绿素含量与抗氧化酶活性的影响[J]. 中南林业科技大学学报, 2017, 37(6): 51-55.
[
|
| [35] |
许春华, 陈全家, 郭忠军, 等. 两种十字花科短命植物抗旱性分析及其指标研究[J]. 新疆农业大学学报, 2012, 35(2): 107-111.
[
|
| [36] |
李海碧, 祝开, 桂意云, 等. 跨代干旱锻炼对甘蔗渗透调节物质、抗氧化酶活性和内源激素含量的影响[J]. 广西糖业, 2023, 43(2): 6-9.
[
|
| [37] |
马新超, 轩正英, 闵昊哲, 等. 水氮耦合对沙培黄瓜叶片抗氧化酶及水氮利用的影响[J]. 节水灌溉, 2022(7): 58-64.
[
|
| [38] |
|
| [39] |
郭玉双, 李祥羽, 任学良. 植物体内活性氧(ROS)的产生及其作用研究进展[J]. 黑龙江农业科学, 2011(8): 146-148.
[
|
| [40] |
朱素琴, 何玲艳, 张千千, 等. 植物细胞内ROS的种类及其产生部位[J]. 安徽农业科学, 2012, 40(34): 16536-16540.
[
|
| [41] |
杨利, 王波, 李文姣, 等. 干旱胁迫下ROS的产生、清除及信号转导研究进展[J]. 生物技术通报, 2021, 37(4): 194-203.
[
|
| [42] |
龚吉蕊, 赵爱芬, 张立新, 等. 干旱胁迫下几种荒漠植物抗氧化能力的比较研究[J]. 西北植物学报, 2004, 24(9): 1570-1577.
[
|
| [43] |
周恒, 谢彦杰. 植物氧化胁迫信号应答的研究进展[J]. 生物技术通报, 2023, 39(11): 36-43.
[
|
| [44] |
田艳丽, 种培芳, 陆文涛, 等. 模拟氮沉降和降水变化对红砂(Reaumuria soongorica)、珍珠猪毛菜(Salsola passerina)生理的影响[J]. 中国沙漠, 2021, 41(3): 165-173.
[
|
| [45] |
祁伟亮, 孙万仓, 马骊. 活性氧参与调控植物生长发育和胁迫应激响应机理的研究进展[J]. 干旱地区农业研究, 2021, 39(3): 69-81, 193.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}