
气候变化背景下白皮松在中国潜在适宜分布预测
|
樊玉科(2000-),男,硕士研究生,主要从事植物多样性研究. E-mail: fanyuke823@foxmail.com |
收稿日期: 2024-04-01
修回日期: 2024-07-13
网络出版日期: 2025-08-12
基金资助
国家自然科学基金项目(31860197)
青藏高原东北边缘云冷杉火烧迹地森林演替动态及演替机理研究(03619078)
Prediction of potential suitable distribution area of Pinus bungeana in China under the background of climate change
Received date: 2024-04-01
Revised date: 2024-07-13
Online published: 2025-08-12
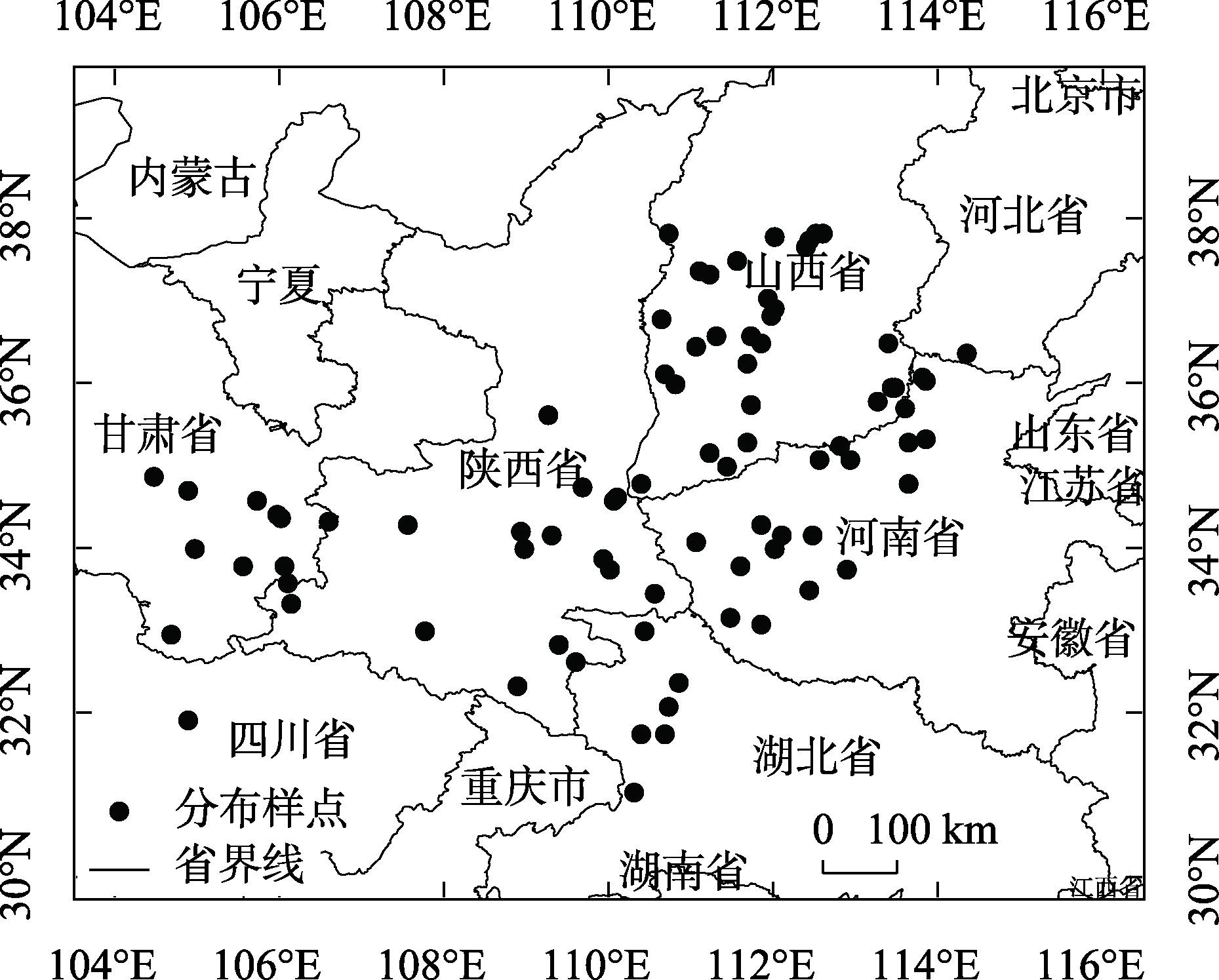
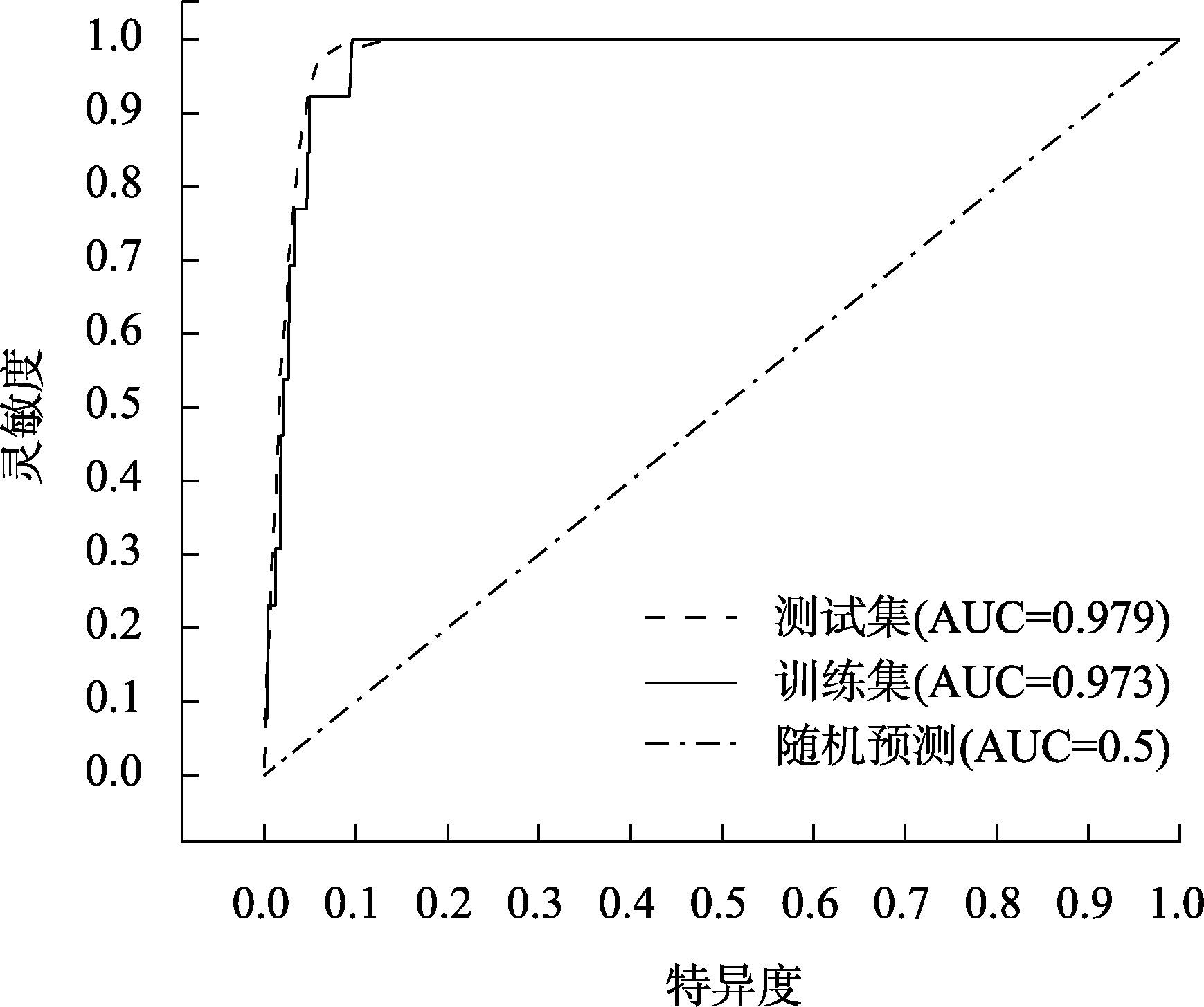
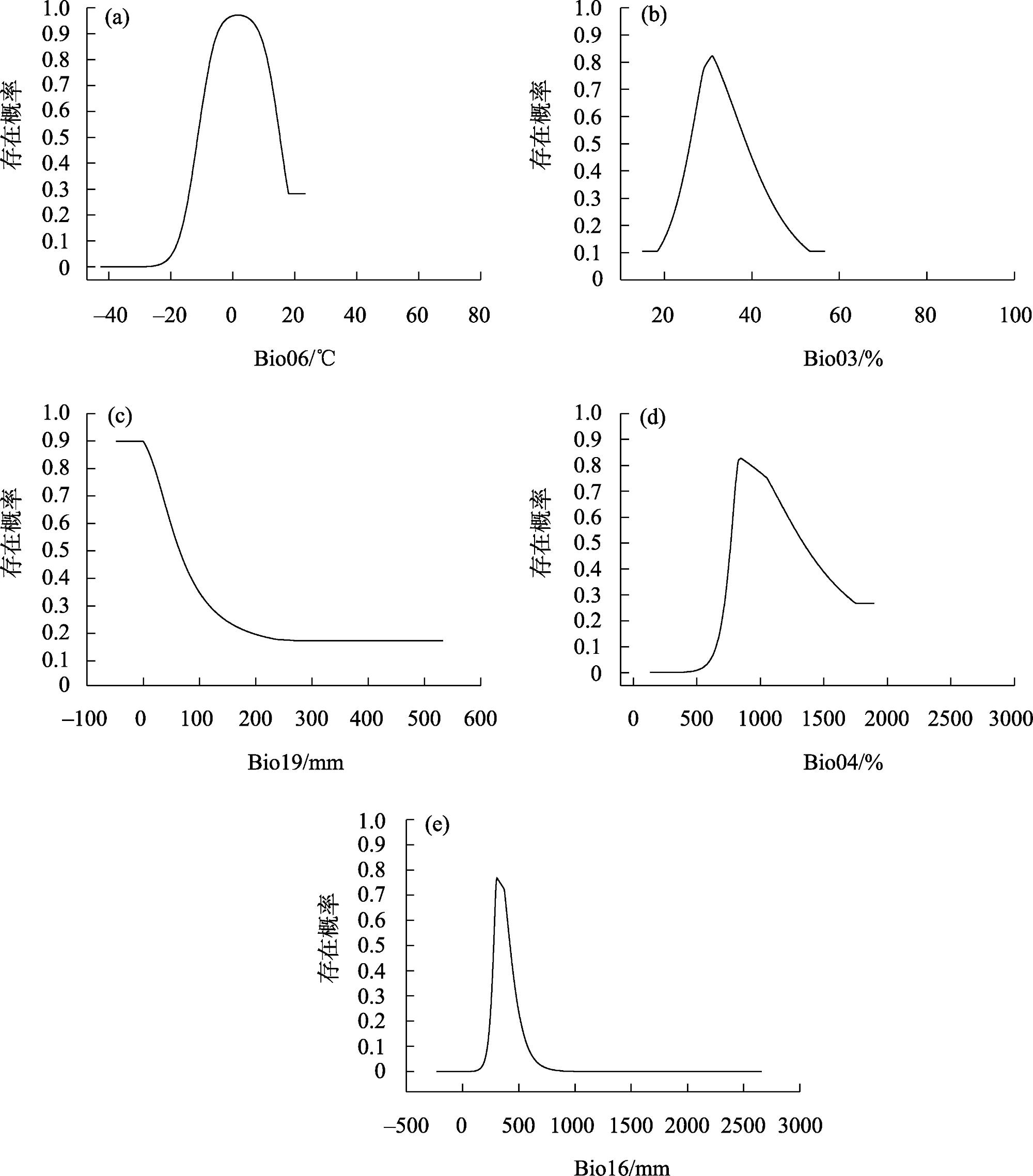
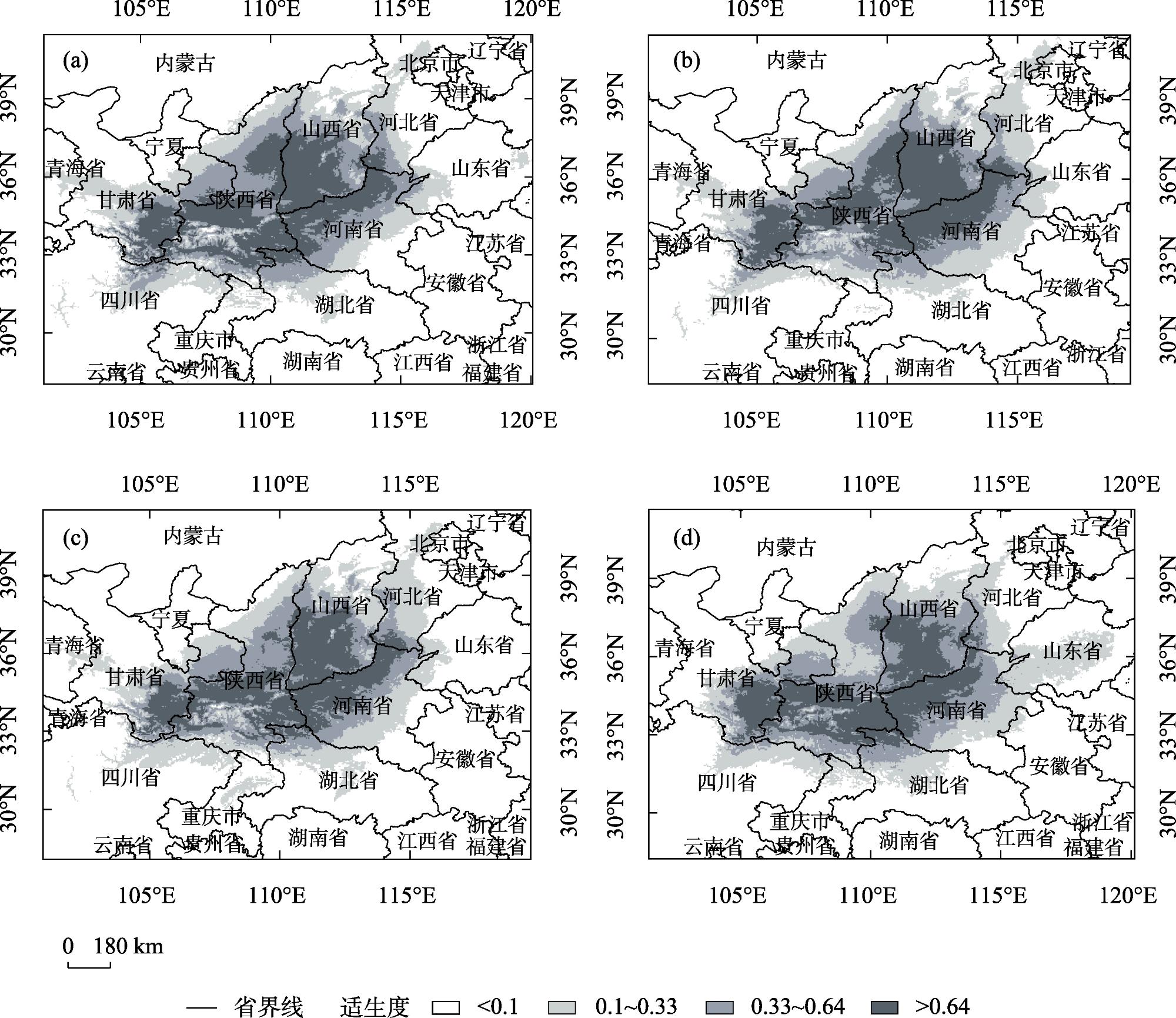
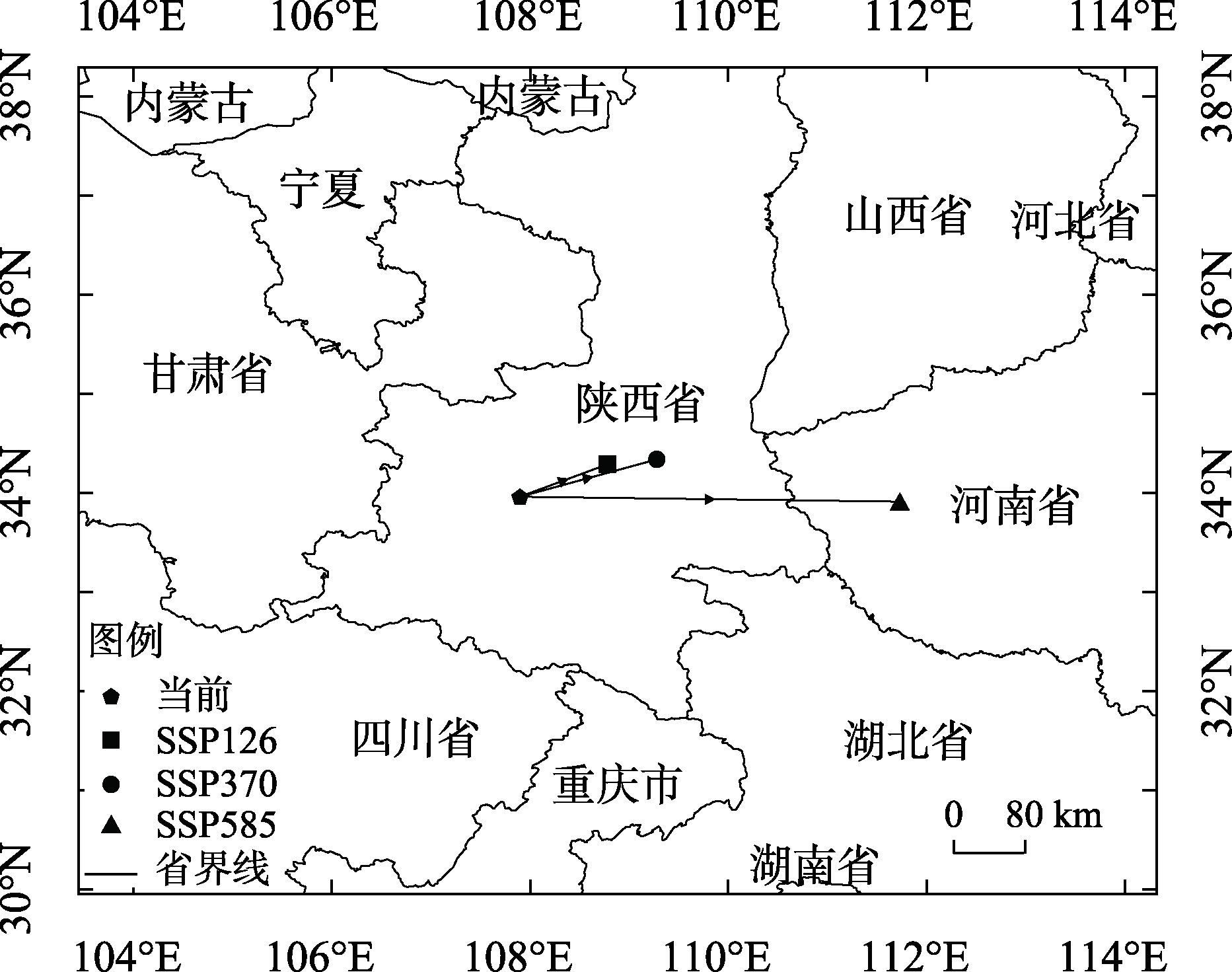
研究旨在预测白皮松潜在适生区分布及气候变化对其的影响,明确其未来适宜的分布范围,为白皮松保护和其在生态工程建设中的应用提供参考。基于白皮松83个野生分布点及气候因子数据,利用MaxEnt模型和ArcGIS软件,模拟白皮松当前和未来3种(2080—2100年,低温室气体排放情景、中温室气体排放情景和高温室气体排放情景)气候变化情景(SSP126、SSP370、SSP585)下的潜在适生区分布。MaxEnt模型模拟结果AUC(受试者工作特征曲线下面积值)值均>0.973,预测结果精度较高。当前气候条件下,白皮松的潜在适生区主要分布在陕西省、山西省南部、甘肃省东南部、河南省和湖北省西北部等地,总面积约74.5×104 km2,未来气候变化背景下,核心适生区面积均有不同程度的缩减,温度为白皮松潜在适生区分布的主限制因子;中、低温室气体排放情景下,白皮松潜在适生区分布的限制因子仍以温度为主;高温室气体排放情景下,全球温度升高加快,降雨量成为影响白皮松适生区分布的主要限制因子,白皮松适生区质心向东偏移,尤其在温室气体高排放浓度下更敏感,迁移距离更远。本文结合白皮松当前和未来潜在适生区的变化,提出了对白皮松保护的建议,且对利用白皮松进行生态工程建设提供参考价值。
关键词: 最大熵(MaxEnt)模型; 白皮松; 气候变化; 潜在分布区; 中国
樊玉科 , 任菊 , 王润龙 , 周栋栋 , 潘自凯 , 张晓玮 , 周晓雷 . 气候变化背景下白皮松在中国潜在适宜分布预测[J]. 干旱区研究, 2024 , 41(10) : 1719 -1730 . DOI: 10.13866/j.azr.2024.10.10
This study was conducted to predict the distribution of potential suitable area and the impact of climate change and to determine the appropriate distribution range in the future, which could provide a reference for the protection of Pinus bungeana and its utilization in ecological engineering construction. Based on 83 wild distribution sites of P. bungeana and climate factor data, the MaxEnt model and ArcGIS software were used to simulate the potential suitable zone distribution of P. bungeana under the present and three climate change scenarios (SSP126, SSP370, and SSP585) (2080-2100, low-level, medium-level, and high greenhouse gas emission scenarios). In the MaxEnt model simulation, the AUC (area value under the subject operating characteristic curve) was >0.973, and the prediction results were highly accurate. Under the present climate conditions, the potential suitable areas of P. bungeana were primarily distributed in Shaanxi Province, southern Shanxi Province, southeastern Gansu Province, northwestern Henan Province, and northwestern Hubei Province, with a total area of approximately 74.5×104 km2, under the background of future climate change. The core suitable distribution areas were reduced to different degrees, with temperature being the primary limiting factor for the distribution of the potential growth zones of P. bungeana. Under low and medium greenhouse gas emission scenarios, temperature still remained the limiting factor for the distribution of the potential growth zones of P. bungeana. Under the high greenhouse gas emission scenario, the global temperature increased faster, and rainfall was the major limiting factor affecting the distribution of the suitable area of P. bungeana. The centroid of the suitable area of P. bungeana shifted eastward, especially being more sensitive under the high emission concentration of greenhouse gases, and the migration distance was farther. This study proposes the protection of P. bungeana, and the results provide a reference for ecological engineering construction using P. bungeana.
表1 气候变量数据Tab. 1 Climate variable data |
| 数据图层编号 | 数据说明 | 数据图层编号 | 数据说明 |
|---|---|---|---|
| Bio01 | 年平均温度/℃ | Bio11 | 最冷季度的平均温度/℃ |
| Bio02 | 每月温差的平均值/℃ | Bio12 | 年降水量/mm |
| Bio03 | 等温性/% | Bio13 | 最湿月的降水量/mm |
| Bio04 | 温度季节性差异/% | Bio14 | 最干月的降水量/mm |
| Bio05 | 最热月的最高温度/℃ | Bio15 | 季节性降水量变异系数 |
| Bio06 | 最冷月的最低温度/℃ | Bio16 | 最湿季度的降水量/mm |
| Bio07 | 温度年较差/℃ | Bio17 | 最干季度的降水量/mm |
| Bio08 | 最湿季度的平均温度/℃ | Bio18 | 最热季度的降水量/mm |
| Bio09 | 最干季度的平均温度/℃ | Bio19 | 最冷季度的降水量/mm |
| Bio10 | 最热季度的平均温度/℃ |
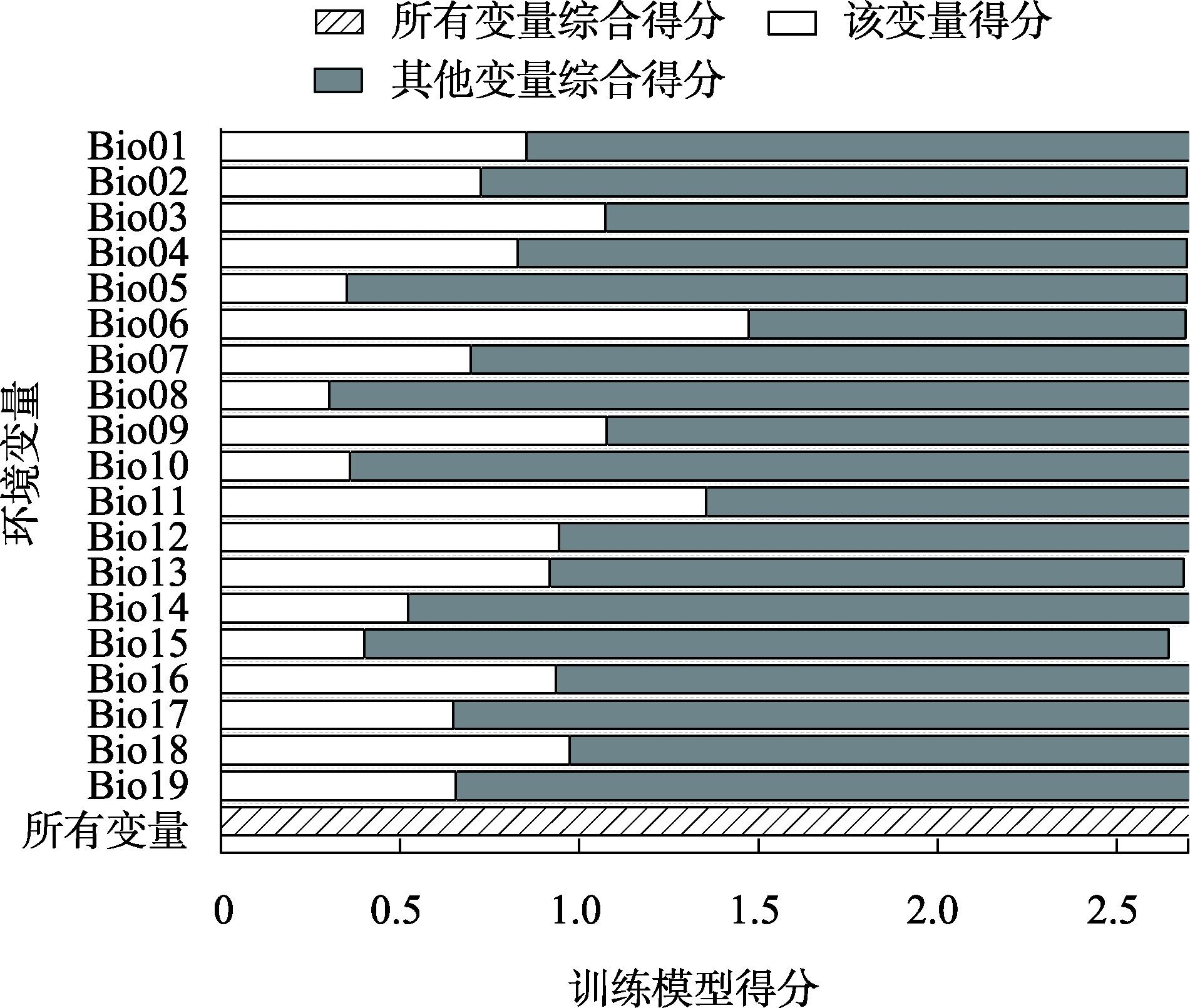
表2 MaxEnt模型中各气候因子对白皮松分布的贡献率Tab. 2 Contribution rate of each climatic factors for the distribution of Pinus bungeana in MaxEnt model |
| 代码 | 当前 | 2080—2100年 | ||
|---|---|---|---|---|
| SSP126 | SSP370 | SSP585 | ||
| Bio06 | 21.1% | 24.5% | 24.1% | 18.9% |
| Bio03 | 16.4% | 9.6% | 10.5% | 19.8% |
| Bio16 | 15.4% | 7.7% | 6.3% | 0.1% |
| Bio19 | 11.6% | 15.1% | 13.7% | 10.5% |
| Bio04 | 9.2% | 14.3% | 14.6% | 12.5% |
| Bio12 | 2% | 13.8% | 14.9% | 21.4% |
表3 未来不同气候情景下白皮松潜在适生区面积Tab. 3 Potential suitable area of Pinus bungeana under different climate scenarios in the future |
| 适生区 | 当前面积 /104 km2 | 2080—2100年SSP126 | 2080—2100年SSP370 | 2080—2100年SSP585 | |||||
|---|---|---|---|---|---|---|---|---|---|
| 面积/104 km2 | 面积变化比例/% | 面积/104 km2 | 面积变化比例/% | 面积/104 km2 | 面积变化比例/% | ||||
| 高适生区 | 23.23 | 19.38 | -16.57 | 20.60 | -11.32 | 20.48 | -11.84 | ||
| 中适生区 | 22.40 | 22.37 | -0.13 | 19.99 | -10.75 | 22.46 | 0. 27 | ||
| 低适生区 | 28.95 | 29.15 | 0.69 | 30.17 | 4.21 | 31.97 | 10.43 | ||
| 总计 | 74.58 | 70.90 | -4.93 | 70.76 | -5.12 | 74.91 | 0.44 | ||
| [1] |
黄蕊, 徐利岗, 刘俊民. 中国西北干旱区气温时空变化特征[J]. 生态学报, 2013, 33(13): 4078-4089.
[
|
| [2] |
黄小燕, 李耀辉, 冯建英, 等. 中国西北地区降水量及极端干早气候变化特征[J]. 生态学报, 2015, 35(5): 1359-1370.
[
|
| [3] |
张雅茜, 王淋, 包福海, 等. 应用最大熵模型预测的欧李潜在适生区分布及气候变化对其的影响[J]. 东北林业大学学报, 2023, 51(11): 54-62.
[
|
| [4] |
|
| [5] |
王卓妮, 袁佳双, 庞博, 等. IPCC AR6 WGⅢ报告减缓主要结论、亮点和启示[J]. 气候变化研究进展, 2022, 18(5): 531-537.
[
|
| [6] |
陈建国, 杨扬, 孙航. 高山植物对全球气候变暖的响应研究进展[J]. 应用与环境生物学报, 2011, 17(3): 435-446.
[
|
| [7] |
王馨, 杨淑桂, 于芬, 等. 檫木的研究进展[J]. 南方林业科学, 2015, 43(5): 29-33, 39.
[
|
| [8] |
|
| [9] |
叶兴状, 张明珠, 赖文峰, 等. 基于MaxEnt优化模型的闽楠潜在适宜分布预测[J]. 生态学报, 2021, 41(20): 8135-8144.
[
|
| [10] |
|
| [11] |
|
| [12] |
魏博, 孙芳芳, 马新, 等. 荒漠濒危植物裸果木适宜分布区对未来气候变化情景的可能响应[J]. 石河子大学学报(自然科学版), 2019, 37(4): 490-497.
[
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
Beaumont Linda J Hughes Lesley,
|
| [17] |
许仲林, 彭焕华, 彭守璋. 物种分布模型的发展及评价方法[J]. 生态学报, 2015, 35(2): 557-567.
[
|
| [18] |
|
| [19] |
|
| [20] |
王晓帆, 段雨萱, 金露露, 等. 基于优化的最大熵模型预测中国高山栎组植物的历史、现状与未来分布变化[J]. 生态学报, 2023, 43(16): 6590-6604.
[
|
| [21] |
|
| [22] |
方文培, 张泽荣. 中国植物志第五十二卷第二分册[M]. 北京: 科学出版社, 1983: 64-65.
[
|
| [23] |
|
| [24] |
彭重华, 薄楠林. 白皮松研究进展[J]. 中国农学通报, 2007, 23(11): 174-178.
[
|
| [25] |
李斌, 顾万春. 白皮松分布特点与研究进展[J]. 林业科学研究, 2003, 16(2): 225-232.
[
|
| [26] |
张建国, 李吉跃, 姜金璞. 京西山区人工林水分参数的研究(Ⅲ)[J]. 北京林业大学学报, 1994, 16(4): 46-54.
[
|
| [27] |
张力强. 白皮松群落结构与生产力研究[D]. 北京: 北京林业大学, 2007.
[
|
| [28] |
岳华峰, 吴晗彬, 樊巍, 等. 白皮松天然林更新与种群结构分析[J]. 中南林业科技大学学报, 2023, 43(3): 99-106, 154.
[
|
| [29] |
赵焱, 张学忠, 王孝安. 白皮松天然林地理分布规律研究[J]. 西北植物学报, 1995, 15(2): 161-166.
[
|
| [30] |
|
| [31] |
邓鑫. 基于Elman神经网络的白皮松潜在适生区估测及未来分布趋势研究[D]. 西安: 西北大学, 2013.
[
|
| [32] |
唐梦, 陈静, 杨灵懿, 等. 气候变化下中国主要生物燃油树种分布与变迁[J]. 生态学报, 2023, 43(24): 10156-10170.
[
|
| [33] |
张华, 赵浩翔, 徐存刚. 气候变化背景下孑遗植物桫椤在中国的潜在地理分布[J]. 生态学杂志, 2021, 40(4): 968-979.
[
|
| [34] |
张艳芳. 基于MaxEnt模型预测花烟草全球潜在适生区的研究[D]. 泰安: 山东农业大学, 2024.
[
|
| [35] |
宋金岳. 基于生态位模型的气候变化情景下红火蚁在中国的适生区预测[D]. 兰州: 西北师范大学, 2022.
[
|
| [36] |
|
| [37] |
曹雪萍, 王婧如, 鲁松松, 等. 气候变化情景下基于最大熵模型的青海云杉潜在分布格局模拟[J]. 生态学报, 2019, 39(14): 5232-5240.
[
|
| [38] |
张晓玮, 王婧如, 王明浩, 等. 中国云杉属树种地理分布格局的主导气候因子[J]. 林业科学, 2020, 56(4): 1-11.
[
|
| [39] |
魏鹏, 张源, 何佳遥, 等. 基于MaxEnt模型分析气候变化下玉米褪绿斑驳病毒的潜在地理分布[J]. 植物保护学报, 2022, 49(5): 1367-1376.
[
|
| [40] |
赖雨曈. CMIP6全球气候模式对中国地区干旱模拟能力评估与未来预估[D]. 北京: 中国气象科学研究院, 2023.
[
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
史柠瑞, 朱珠, 王艳莉. 基于MaxEnt模型对2种青兰属植物在未来气候变化下潜在分布的预测研究[J]. 中国农学通报, 2023, 39(32): 115-123.
[
|
| [45] |
宦智群, 耿兴敏, 徐小蓉, 等. 基于MaxEnt模型分析不同气候变化情景下的黄心夜合(Michelia martinii)潜在地理分布[J]. 生态与农村环境学报, 2023, 39(10): 1277-1287.
[
|
| [46] |
李晓辰, 贡璐, 魏博, 等. 气候变化对新疆雪岭云杉潜在适宜分布及生态位分化的影响[J]. 生态学报, 2022, 42(10): 4091-4100.
[
|
| [47] |
赵儒楠, 何倩倩, 褚晓洁, 等. 气候变化下千金榆在我国潜在分布区预测[J]. 应用生态学报, 2019, 30(11): 3833-3843.
[
|
| [48] |
熊中人, 张晓晨, 邹旭, 等. 中国天山花楸适生区预测及其对气候变化的响应[J]. 生态科学, 2019, 38(5): 44-51.
[
|
| [49] |
刘维, 赵儒楠, 圣倩倩, 等. 矮牡丹在中国的地理分布及潜在分布区预测[J]. 北京林业大学学报, 2021, 43(12): 83-92.
[
|
| [50] |
欧阳林男, 陈少雄, 张维耀, 等. 柠檬桉在中国的适生地理分布及其影响因子[J]. 生态学杂志, 2019, 38(2): 361-367.
[
|
| [51] |
钟淇涵. 基于物种分布模型的科尔沁沙地元宝枫潜在适生区研究[D]. 呼和浩特: 内蒙古农业大学, 2023.
[
|
| [52] |
|
| [53] |
应邦肯, 田阔, 郭浩宇, 等. 基于MaxEnt模型预测未来气候变化情境下红树秋茄(Kandelia obovata)在中国潜在适生区的变化[J]. 生态学报, 2024, 44(1): 224-234.
[
|
| [54] |
张丹红, 王效科, 张路, 等. 大比例尺土壤保持服务制图分级方法研究[J]. 生态学报, 2021, 41(4): 1391-1401.
[
|
| [55] |
|
| [56] |
李林霞, 何兰君, 席磊, 等. 中国南方松林地理替代分布规律及其气候主导因子研究[J]. 西南林业大学学报(自然科学), 2024, 44(1): 97-105.
[
|
| [57] |
张雷, 王琳琳, 刘世荣, 等. 生境概率预测值转换为二元值过程中4个阈值选择方法的比较评估——以珙桐和杉木生境预估为例[J]. 植物生态学报, 2017, 41(4): 387-395.
[
|
| [58] |
|
| [59] |
|
| [60] |
朱飙. 西北地区气候暖湿化背景下水汽、潜在蒸散及极端温度和降水的变化特征[D]. 兰州: 兰州大学, 2023.
[
|
| [61] |
梁玉莲, 延晓冬. RCPs情景下中国21世纪气候变化预估及不确定性分析[J]. 热带气象学报, 2016, 32(2): 183-192.
[
|
| [62] |
翟颖佳. 中国华北地区和西北东部干旱气候变化特征[D]. 兰州: 兰州大学, 2014.
[
|
| [63] |
郭聪聪, 沈永宝, 史锋厚. 白皮松种子休眠研究进展[J]. 南京林业大学学报(自然科学版), 2019, 43(2): 175-183.
[
|
| [64] |
郭聪聪, 沈永宝, 史锋厚. 白皮松种子萌发过程中呼吸代谢和内源激素对温度变化的响应[J]. 中南林业科技大学学报, 2021, 41(3): 25-36.
[
|
| [65] |
郭聪聪, 沈永宝, 史锋厚. 温度对白皮松种子萌发过程中储藏物质代谢及酶活性的影响[J]. 南京林业大学学报(自然科学版), 2023, 47(6): 25-34.
[
|
| [66] |
彭剑峰, 刘玉振, 王婷. 神农山白皮松不同龄组年轮—气候关系及PDSI重建[J]. 生态学报, 2014, 34(13): 3509-3518.
[
|
| [67] |
|
| [68] |
|
| [69] |
欧阳林男, 陈少雄, 刘学锋, 等. 赤桉在中国的适生地理区域及其对气候变化的响应[J]. 林业科学, 2019, 55(12): 1-11.
[
|
| [70] |
李宁宁, 张爱平, 张林, 等. 气候变化下青藏高原两种云杉植物的潜在适生区预测[J]. 植物研究, 2019, 39(3): 395-406.
[
|
| [71] |
张晓玮, 蒋玉梅, 毕阳, 等. 基于MaxEnt模型的中国沙棘潜在适宜分布区分析[J]. 生态学报, 2022, 42(4): 1420-1428.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}