
西北干旱区藓类结皮覆盖下土壤多功能性特征及影响因子
|
雷菲亚(1997-),女,硕士研究生,主要从事干旱区极端环境对藓类结皮的影响. E-mail: 2683138732@qq.com |
收稿日期: 2024-02-06
修回日期: 2024-03-17
网络出版日期: 2024-06-20
基金资助
新疆天山英才项目(2022TSYCCX0001)
新疆天山英才项目(2022TSYCLJ0058)
国家自然科学基金(U2003214)
国家自然科学基金(42377358)
Characterization of soil multifunctionality and its determining factors under moss crust cover in the arid regions of Northwest China
Received date: 2024-02-06
Revised date: 2024-03-17
Online published: 2024-06-20
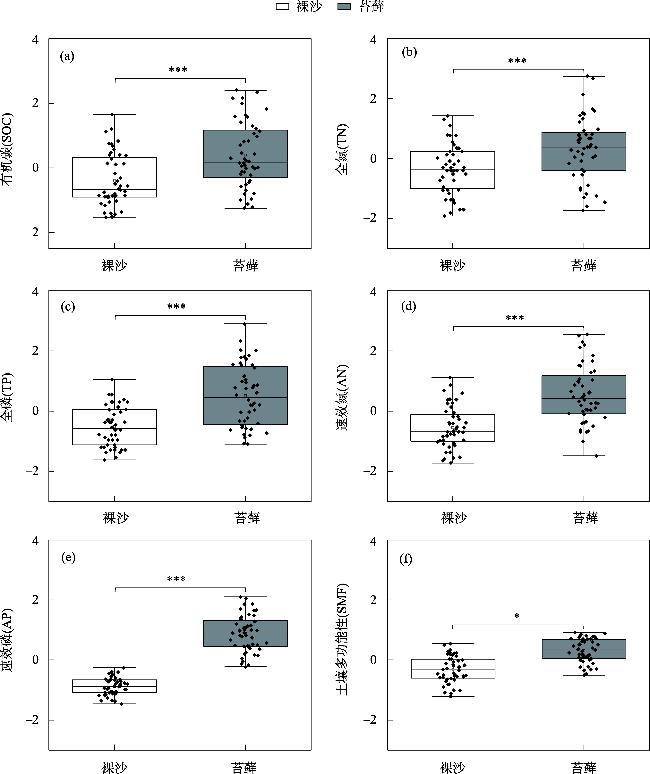
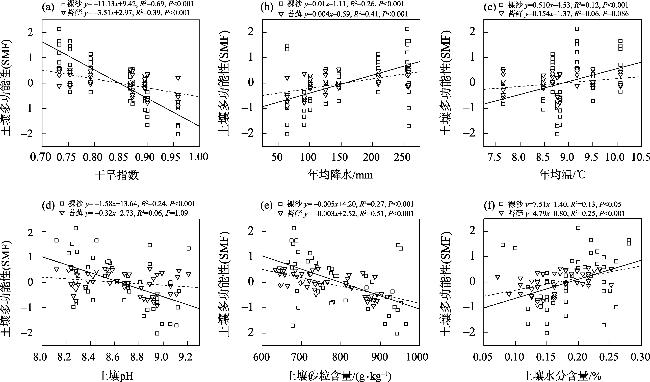
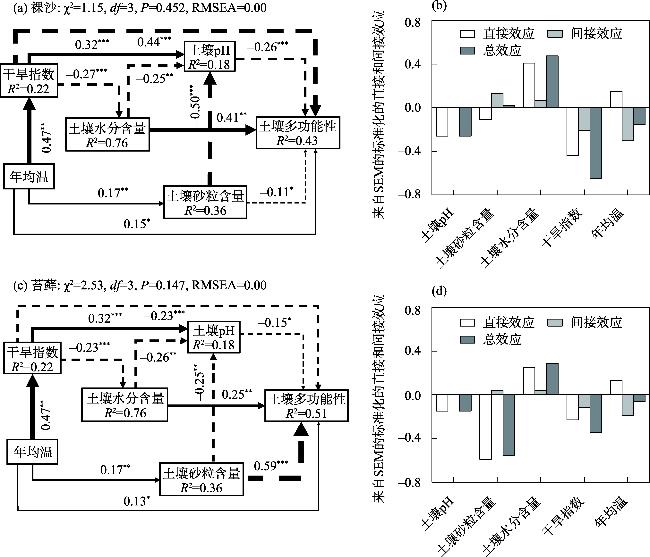
生物土壤结皮是由干旱区重要的活性地被物组成,可显著影响地表土壤的物质循环与能量交换,改善表层土壤的物理、化学和生物学性质,从而影响土壤多功能性(Soil Multifunctionality,SMF)。藓类结皮是生物土壤结皮的重要类型之一,本研究旨在探究西北干旱区荒漠藓类结皮覆盖下土壤与裸沙的SMF差异性,探究两者SMF变化的主要驱动因素。通过分析土壤的8个干旱区关键生态系统功能指标,运用平均值法和因子分析法计算SMF,运用最小二乘回归分析和结构方程模型(SEM)等探究SMF变化的主要驱动因素。研究发现:(1) 藓类结皮覆盖下土壤的单一和多功能性显著高于裸沙。(2) 裸沙和藓类结皮覆盖下SMF变化的驱动要素具有差异性,裸沙SMF的主要驱动因素为干旱(Aridity)和土壤含水量(SWC),而藓类结皮覆盖下SMF驱动要素为土壤砂粒含量。(3) 年均温(MAT)对裸沙和藓结皮覆盖土壤SMF的变化均呈现最大的间接效应。因此,藓类结皮发育显著增加了荒漠土壤SMF,同时也调节SMF的相关驱动因素。以上研究结果对深入理解荒漠裸沙和结皮覆盖下SMF的差异性及驱动因素具有重要意义。
雷菲亚 , 李小双 , 陶冶 , 尹本丰 , 荣晓莹 , 张静 , 陆永兴 , 郭星 , 周晓兵 , 张元明 . 西北干旱区藓类结皮覆盖下土壤多功能性特征及影响因子[J]. 干旱区研究, 2024 , 41(5) : 812 -820 . DOI: 10.13866/j.azr.2024.05.09
Biological soil crusts (BSCs) are the main active groundcover community in arid regions. BSCs can significantly affect the material cycle and energy exchange, improve the physical, chemical, and biological properties of surface soil, and influence the soil multifunctionality (SMF). Moss crust is an important type of BSCs. This study investigates the SMF variability of moss crust-covered and bare sand in the deserts of northwestern arid regions, and explored the main drivers of the variability. We analyzed eight crucial ecosystem function indicators. SMF was calculated by applying the mean method and factor analysis approach. We used the ordinary least square and structural equation modeling to explore the drivers of SMF changes. The results show that: (1) soil monofunctionality and SMF under moss crust cover were higher than those in bare sand (P<0.05). (2) The drivers of the SMF change in bare sand and under moss crust cover were very different. The main drivers of SMF in bare sand were aridity and soil water content, whereas the driver of SMF under moss crust cover was soil sand content (Sand). (3) The mean annual temperature had the largest indirect effect on changes in SMF for both soil in bare sand and under moss crust cover. Therefore, the development of moss crust significantly increased SMF and, in addition, modulated the relevant drivers of SMF. Our results are important for a deep understanding of the differences and drivers of SMF in desert soil with bare sand and under moss crust cover.
表1 8个采样点的地理、植被、土壤和气候信息Tab. 1 Geographic, vegetation, soil and climatic information of the eight sampling sites |
| 样点 | 经度/(°) | 纬度/(°) | 海拔/m | 植被盖度 | 土壤类型 | 结皮盖度 | 结皮类型 | 土壤粒级分布/% | 年均 降水/mm | 年均 气温/℃ | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 粉粒 | 砂粒 | 石砾 | ||||||||||
| S1 | 87.15417 | 45.3023 | 461.79 | 0.05 | 简育砂性土 | 0.74 | 藻-地-藓 | 4.94 | 67.09 | 27.98 | 91.50 | 8.82 |
| S2 | 87.67692 | 45.2478 | 575.22 | 0.01 | 简育砂性土 | 0.84 | 藻-地-藓 | 4.61 | 83.08 | 12.31 | 101.03 | 8.48 |
| S3 | 88.30511 | 45.2636 | 729.83 | 0.04 | 简育砂性土 | 0.85 | 藻-地-藓 | 1.94 | 95.25 | 2.81 | 125.50 | 7.65 |
| S4 | 108.5659 | 38.8198 | 1391.35 | 0.18 | 钙积潜育土 | 0.43 | 藻-地-藓 | 2.03 | 91.14 | 6.83 | 209.00 | 8.67 |
| S5 | 110.2118 | 39.0136 | 1250.04 | 0.01 | 简育砂性土 | 0.45 | 藻-地-藓 | 0.93 | 81.03 | 18.04 | 255.00 | 9.18 |
| S6 | 103.8205 | 40.8864 | 1332.37 | 0.02 | 简育石膏土 | <0.10 | 藻-地-藓 | 1.78 | 82.66 | 15.56 | 64.50 | 8.77 |
| S7 | 108.9267 | 37.9791 | 1158.39 | 0.01 | 过渡性红砂土 | <0.10 | 藻-地-藓 | 3.42 | 75.13 | 21.45 | 257.00 | 10.07 |
| S8 | 104.8801 | 37.5025 | 1508.13 | 0.04 | 简育砂性土 | 0.23 | 藻-地-藓 | 0.97 | 88.95 | 10.07 | 146.15 | 9.50 |
注:藻为藻类结皮;地为地衣结皮;藓为藓类结皮。 |
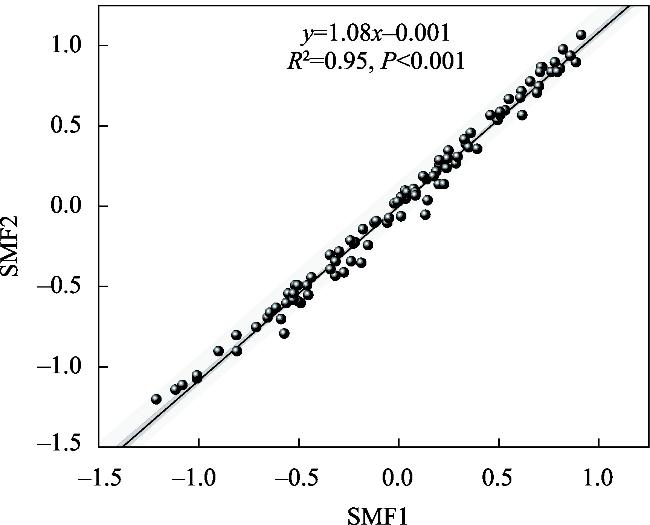
图2 基于因子分析法(SMF1)和平均值法(SMF2)的土壤多功能指数之间的线性关系Fig. 2 Linear relationship of the soil multifunctional index obtained by factor analysis approach(SMF1) and mean method (SMF2) |
表2 土壤单一和多功能的相关性Tab. 2 Correlation between soil mono- and multifunctionality |
| SOC | TN | TP | C:N | C:P | N:P | AN | AP | SMF1 | SMF2 | |
|---|---|---|---|---|---|---|---|---|---|---|
| SMF1 | 0.832** | 0.701** | 0.614** | 0.240* | 0.293** | 0.218* | 0.784** | 0.752** | 1 | 0.994** |
| SMF2 | 0.860** | 0.688** | 0.660** | 0.262** | 0.256* | -0.254* | 0.796** | 0.762** | 0.994** | 1 |
注:SOC表示有机碳,TN表示全氮,TP表示全磷,AN表示速效氮,AP表示速效磷,C:N表示碳氮比,C:P表示碳磷比,N:P表示氮磷比;SMF1表示因子分析法,SMF2表示平均值法;*表示P<0.05;**表示P<0.01;***表示P<0.001。 |
图5 环境因素对裸沙(a~b)和藓类结皮覆盖下(c~d)SMF直接和间接影响的结构方程模型注:实线箭头表示正效应,虚线箭头表示负效应。箭头的粗细与标准化路径系数的大小成正比。*表示P<0.05;**表示P<0.01;***表示P<0.001。χ2为卡方值;df为自由度;RMSEA为均方根近似误差。 Fig. 5 Structural equation modeling for assessment of the direct and indirect effects of environmental factors on SMF in bare sand (a-b) and under moss crusts cover (b-d) |
感谢荒漠与绿洲生态国家重点实验室/干旱区生态安全与可持续发展重点实验室团队人员。
| [1] |
|
| [2] |
蒙文萍, 戴全厚, 冉景丞. 藓植物岩溶作用研究进展[J]. 植物生态学报, 2019, 43(5): 396-407.
[
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
张雨虹, 张韶阳, 张树煇, 等. 毛乌素沙地藓类结皮对沙化土壤性质和细菌群落的影响[J]. 土壤学报, 2021, 58(6): 1585-1597.
[
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
张世航, 陶冶, 陈玉森, 等. 准噶尔荒漠土壤多功能性的空间变异特征及其驱动因素[J]. 生物多样性, 2022, 30(8): 140-150.
[
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
陈亚宁, 杨青, 罗毅, 等. 西北干旱区水资源问题研究思考[J]. 干旱区地理, 2012, 35(1): 1-9.
[
|
| [24] |
郭泽呈, 魏伟, 石培基, 等. 中国西北干旱区土地沙漠化敏感性时空格局[J]. 地理学报, 2020, 75(9): 1949-1965.
[
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
陶冶, 刘耀斌, 吴甘霖, 等. 准噶尔荒漠区域尺度浅层土壤化学计量特征及其空间分布格局[J]. 草业学报, 2016, 25(7): 13-23.
[
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
高丽倩, 赵允格, 许明祥, 等. 生物土壤结皮演替对土壤生态化学计量特征的影响[J]. 生态学报, 2018, 38(2): 678-688.
[
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
李宁宁, 张光辉, 王浩, 等. 黄土丘陵沟壑区生物结皮对土壤抗蚀性能的影响[J]. 中国水土保持科学, 2020, 18(1): 42-48.
[
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}