
Distribution characteristics and its influencing factors of surface soil pollen in the marginal monsoon region of China
Received date: 2024-01-26
Revised date: 2024-05-30
Online published: 2026-03-11
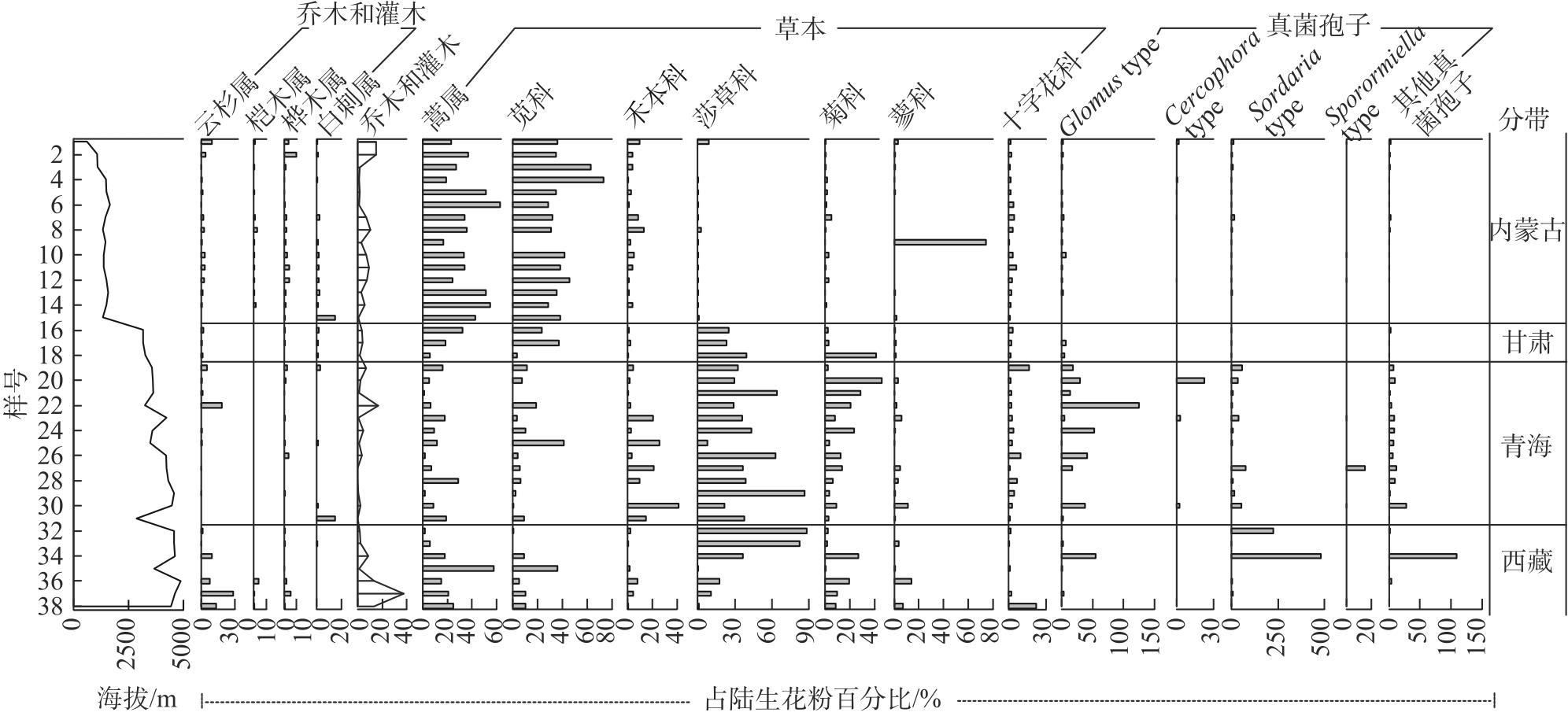
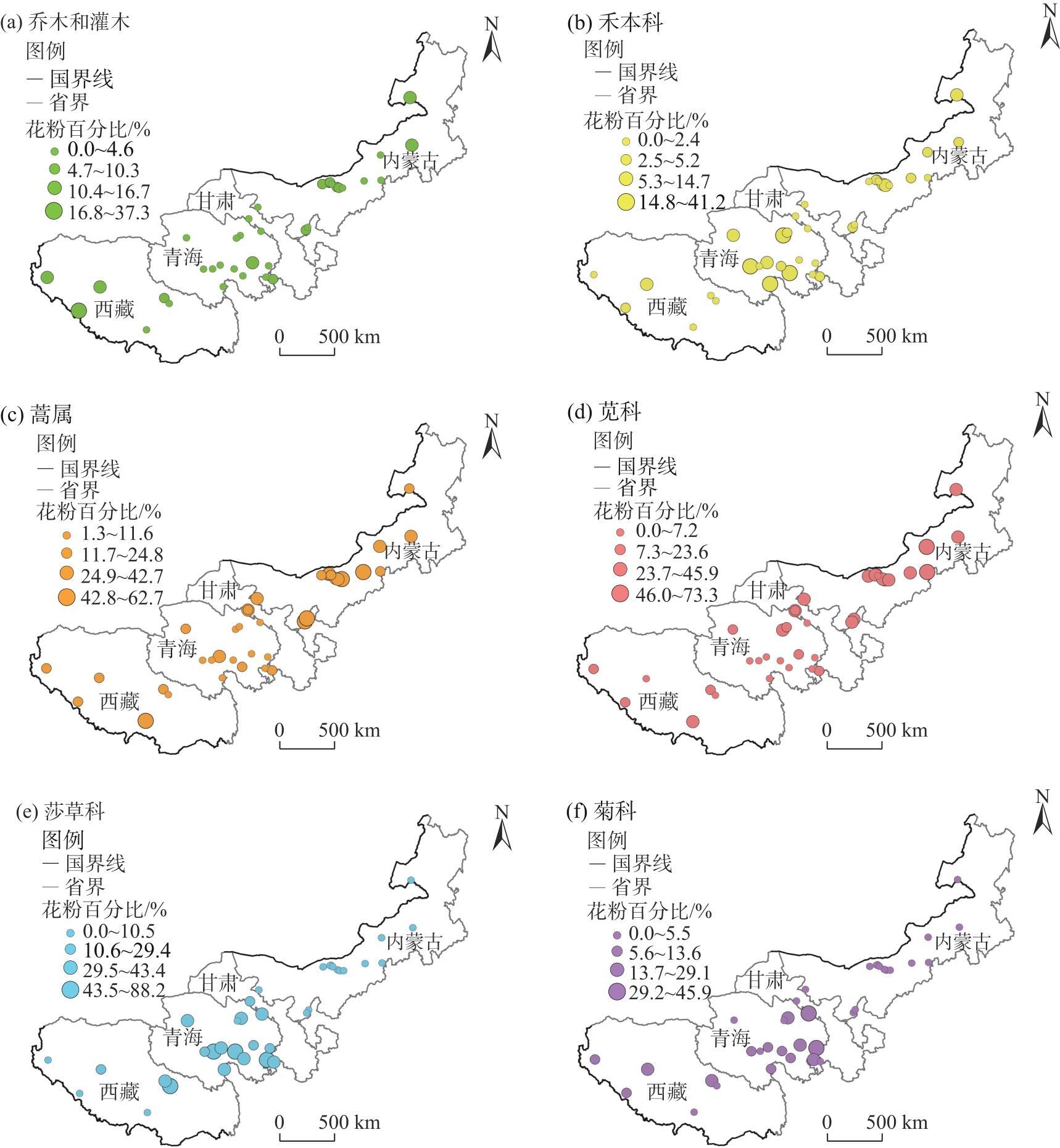
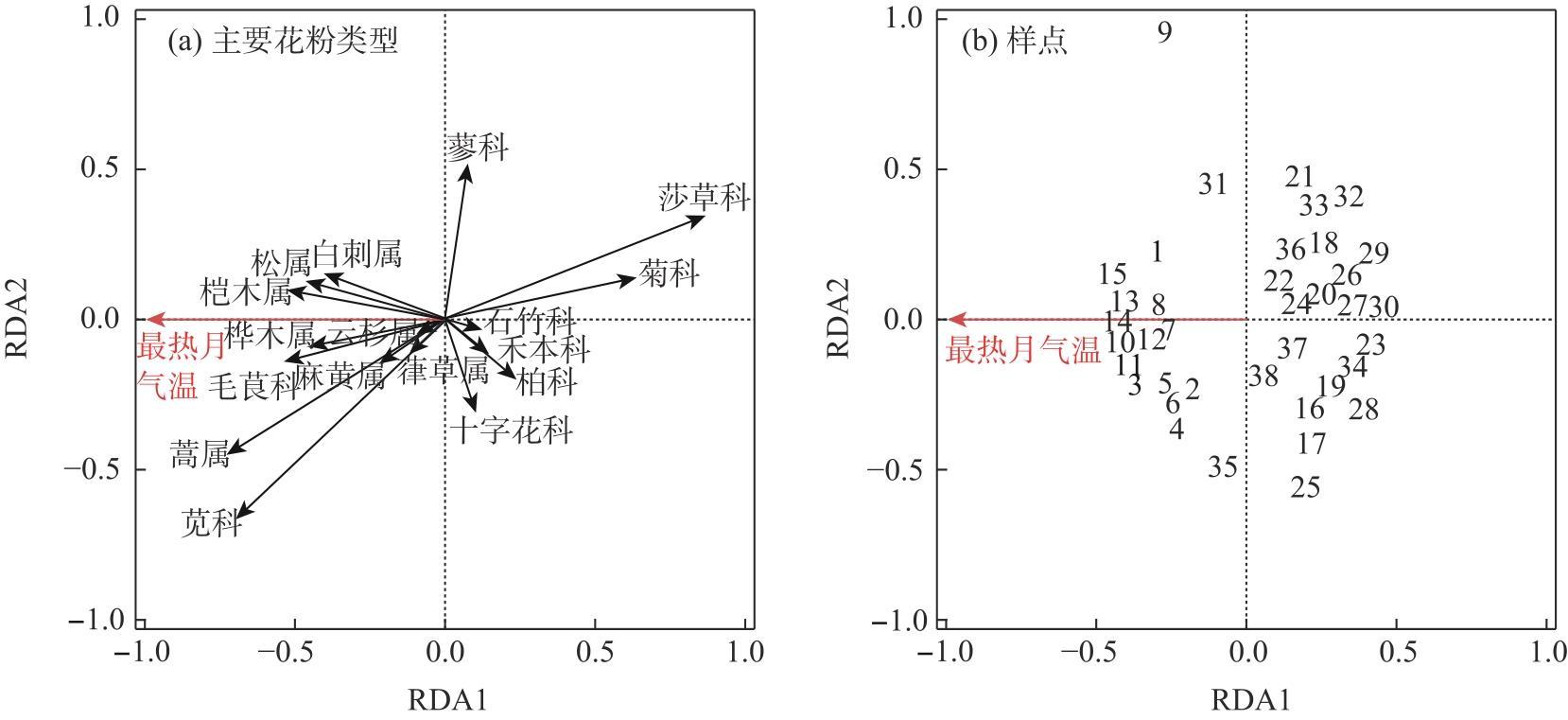
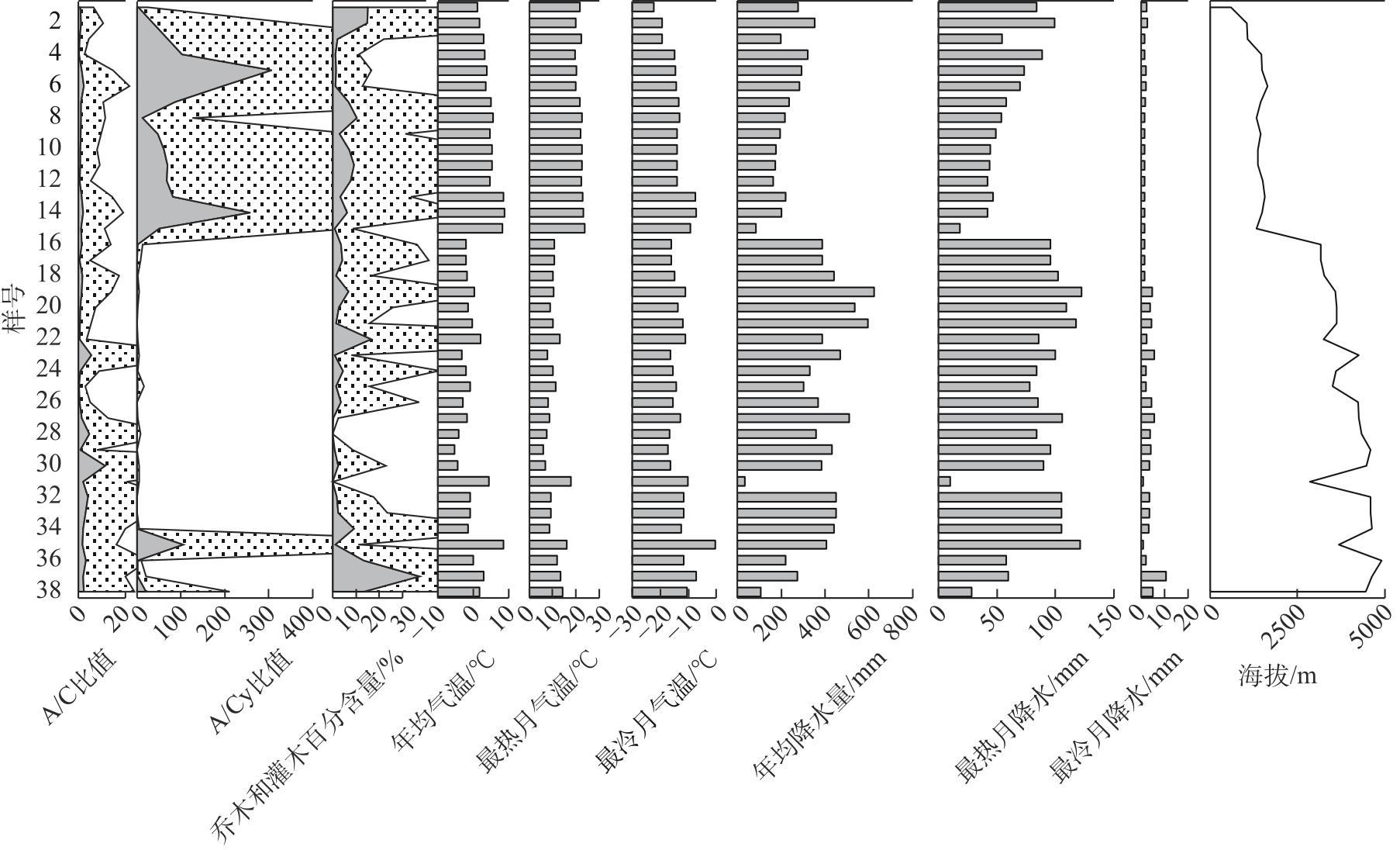
Studying the distribution characteristics of surface soil pollen in the marginal monsoon region of China and the factors influencing this distribution improves the reliability of reconstructing regional vegetation and paleo-monsoon evolution based on fossil pollen data. Through surface soil pollen analysis of 38 sampling sites with annual precipitation ranging from 30-650 mm in the marginal monsoon region of China, this study investigates the relationships among surface pollen assemblages, climate, and human activities. The results reveal that: (1) In low-altitude temperate steppe regions with a relatively warm and dry climate, Amaranthaceae and Artemisia dominate the pollen assemblages, and the ratio of Artemisia/Chenopodiaceae (A/C ratio) generally aligns with average annual precipitation. (2) In high-altitude alpine steppe and meadow regions with a relatively cold and humid climate, pollen assemblages are dominated by Cyperaceae, Asteraceae, and Poaceae. The summed percentages of Artemisia and Amaranthaceae pollen average 25.8%, and no distinctive correlation exists between the A/C ratio and average annual precipitation in these regions. (3) The total tree and shrub pollen percentages exhibit a stronger correlation with average annual temperature and the temperature of the warmest month compared to average annual precipitation. (4) These pollen assemblage characteristics reflect the vegetation distribution and hydrothermal conditions at varying altitudes in the marginal monsoon region of China. Redundancy analysis (RDA) of the pollen data also reveals the distribution of pollen assemblages and major pollen types along gradients of the warmest month temperature and altitude. In addition, the high abundance of fungal spores on the soil surface in the alpine region reflects environmental characteristics associated with significant grazing intensity and a subsequent high soil erosion rate.
Key words: marginal monsoon region; surface soil pollen; vegetation; climate; grazing intensity
Siqi WU , Haiqin WEI , Chunzhu CHEN , Lisi WEI , Wenwei ZHAO , Huan LI , Yi SUN , Qingfeng JIANG , Xiaojian ZHANG , Yan ZHAO . Distribution characteristics and its influencing factors of surface soil pollen in the marginal monsoon region of China[J]. Arid Land Geography, 2025 , 48(1) : 53 -62 . DOI: 10.12118/j.issn.1000-6060.2024.059
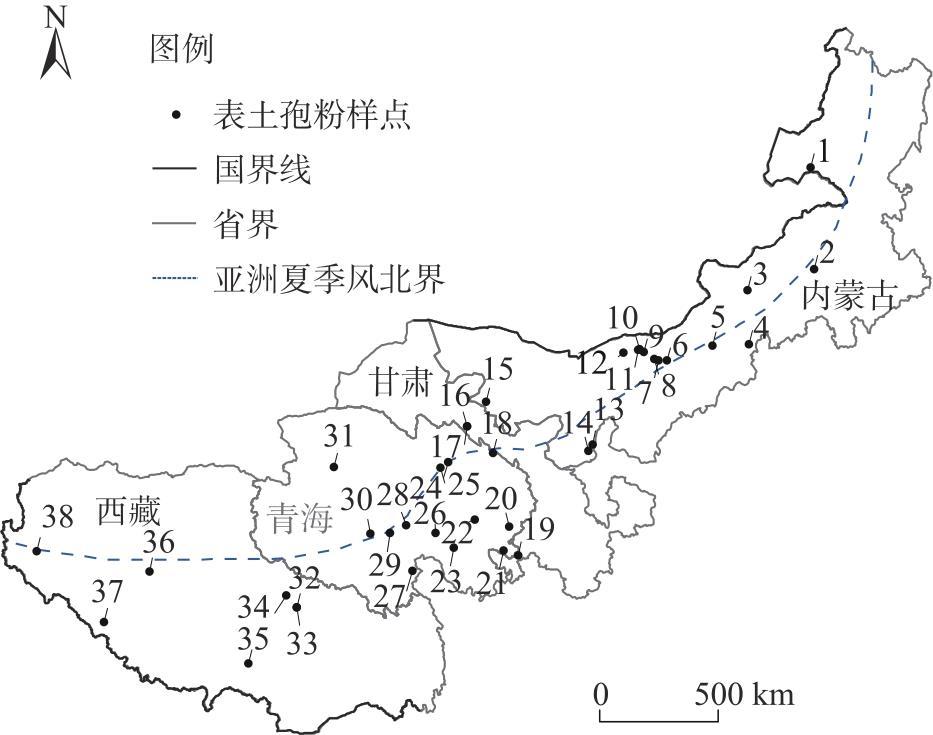
表1 中国季风边缘区38个表土孢粉样品采集信息Tab. 1 Information of 38 surface soil pollen samples in the marginal monsoon region of China |
| 样号 | 省份 | 经度/°E | 纬度/°N | 海拔/m | 植被类型 | 样号 | 省份 | 经度/°E | 纬度/°N | 海拔/m | 植被类型 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 内蒙古 | 118.1916 | 48.1954 | 594 | 温带草原 | 20 | 青海 | 101.7496 | 35.2380 | 3620 | 高寒草甸 |
| 2 | 内蒙古 | 117.6678 | 44.3608 | 1061 | 温带草原 | 21 | 青海 | 101.5095 | 34.3277 | 3624 | 高寒草甸 |
| 3 | 内蒙古 | 114.0218 | 43.8385 | 1079 | 温带草原 | 22 | 青海 | 100.0912 | 35.4433 | 3251 | 高寒草甸 |
| 4 | 内蒙古 | 113.8248 | 41.8171 | 1474 | 温带草原 | 23 | 青海 | 99.1968 | 34.3466 | 4246 | 高寒草原 |
| 5 | 内蒙古 | 111.9580 | 41.8743 | 1484 | 温带草原 | 24 | 青海 | 98.6818 | 37.5435 | 3595 | 高寒草甸 |
| 6 | 内蒙古 | 109.5486 | 41.4184 | 1643 | 温带草原 | 25 | 青海 | 98.3203 | 37.3161 | 3498 | 高寒草原 |
| 7 | 内蒙古 | 109.1272 | 41.4311 | 1442 | 温带草原 | 26 | 青海 | 98.2959 | 34.8516 | 4224 | 高寒草甸 |
| 8 | 内蒙古 | 108.9133 | 41.4994 | 1338 | 温带草原 | 27 | 青海 | 97.3486 | 33.3814 | 4249 | 高寒草原 |
| 9 | 内蒙古 | 108.3732 | 41.7582 | 1451 | 温带草原 | 28 | 青海 | 96.8674 | 35.0698 | 4325 | 高寒草甸 |
| 10 | 内蒙古 | 108.1852 | 41.8837 | 1362 | 温带草原 | 29 | 青海 | 96.1393 | 34.7171 | 4579 | 高寒草甸 |
| 11 | 内蒙古 | 108.0843 | 41.8669 | 1372 | 温带草原 | 30 | 青海 | 95.2436 | 34.6446 | 4472 | 高寒草原 |
| 12 | 内蒙古 | 107.3315 | 41.7723 | 1498 | 温带草原 | 31 | 青海 | 93.1708 | 36.9647 | 2857 | 高寒荒漠 |
| 13 | 内蒙古 | 105.7014 | 38.3493 | 1563 | 温带草原 | 32 | 西藏 | 92.2592 | 31.5912 | 4591 | 高寒草甸 |
| 14 | 内蒙古 | 105.5002 | 38.1114 | 1487 | 温带草原 | 33 | 西藏 | 92.2586 | 31.5915 | 4590 | 高寒草甸 |
| 15 | 内蒙古 | 100.3784 | 39.8783 | 1325 | 温带荒漠 | 34 | 西藏 | 91.7169 | 31.9666 | 4623 | 高寒草甸 |
| 16 | 甘肃 | 99.4784 | 38.9213 | 3172 | 高寒草原 | 35 | 西藏 | 90.4644 | 29.2524 | 3667 | 高寒草原 |
| 17 | 甘肃 | 99.4784 | 38.9214 | 3170 | 高寒草原 | 36 | 西藏 | 85.4007 | 32.0089 | 4890 | 高寒草原 |
| 18 | 甘肃 | 100.8250 | 37.9754 | 3266 | 高寒灌丛草甸 | 37 | 西藏 | 83.8801 | 29.7957 | 4628 | 高寒草原 |
| 19 | 青海 | 102.1994 | 34.1744 | 3569 | 高寒灌木草甸 | 38 | 西藏 | 80.2225 | 31.8252 | 4449 | 高寒草原 |
表2 中国季风边缘区38个表土孢粉样品及气候参数的冗余分析(RDA)结果Tab. 2 RDA results of 38 surface soil pollen samples and the climate variables in the marginal monsoon region of China |
| 气候参数 | 方差膨胀 因子(VIF) | 气候变量作为 唯一预测因子 | 约束轴置换 检验(ANOVA) | ||||
|---|---|---|---|---|---|---|---|
| 运行1 | 运行2 | 方差解释量/% | 方差解释量/% | P值 | |||
| 年均降水量 | 17.3 | 15.8 | 7.2 | 1.0 | 0.129 | ||
| 最热月降水量 | 14.0 | 14.0 | 5.2 | 1.1 | 0.089 | ||
| 最冷月降水量 | 1.8 | 1.6 | 4.0 | 0.8 | 0.158 | ||
| 年均气温 | 446.1 | - | - | - | - | ||
| 最热月气温 | 340.8 | 2.5 | 12.9 | 5.3 | 0.001 | ||
| 最冷月气温 | 97.2 | 1.0 | 0.5 | 0.4 | 0.558 | ||
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
韩岳婷, 李建勇, 刘剑波, 等. 准噶尔盆地西部花粉对植被的指示性研究[J]. 干旱区地理, 2023, 46(5): 773-781.
[
|
| [13] |
|
| [14] |
陈辉, 吕新苗, 李双成. 柴达木盆地东部表土花粉分析[J]. 地理研究, 2004, 23(2): 201-210.
[
|
| [15] |
李芙蓉. 中国北方表土孢粉组合及其与植被和气候的关系[D]. 兰州: 兰州大学, 2012.
[
|
| [16] |
|
| [17] |
|
| [18] |
魏海成, 郑卓, 马海州, 等. 青海表土花粉分布规律及其与植被的关系[J]. 干旱区地理, 2009, 32(6): 932-940.
[
|
| [19] |
程波, 陈发虎. 西北干旱区石羊河流域表土花粉分析[J]. 中国沙漠, 2010, 30(2): 350-356.
[
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
刘鸿雁, 李宜垠. 半干旱区气候变化和人类活动的孢粉指示[J]. 古生物学报, 2009, 48(2): 211-221.
[
|
| [26] |
张雅平, 赵克良, 周新郢, 等. 家养食草动物粪便中的花粉及菌孢子类型及其对人类活动的指示意义[J]. 人类学学报, 2021, 40(5): 879-887.
[
|
| [27] |
郝秀东, 翁成郁. 粪生真菌孢子在古生态学研究中的指示意义[J]. 海洋地质与第四纪地质, 2015, 35(1): 175-184.
[
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
中国植被编辑委员会. 中国植被[M]. 北京: 科学出版社, 1995: 917-1034.
[Editorial Committee of Vegetation of China. Vegetation of China[M]. Beijing: Science Press, 1995: 917-1034.]
|
| [35] |
|
| [36] |
|
| [37] |
王伏雄, 钱南芬, 张玉, 等. 中国植物花粉形态[M]. 北京: 科学出版社, 1995.
[
|
| [38] |
唐领余, 毛礼米, 李春海, 等. 中国第四纪孢粉图鉴[M]. 北京: 科学出版社, 2016.
[
|
| [39] |
席以珍, 宁建长. 中国干旱半干旱地区花粉形态研究[J]. 玉山生物学报, 1994, 11: 119-191.
[
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}