
Distribution characteristics and driving factors of soil fungi between alfalfa roots in different habitats in northern Tianshan Mountains
Received date: 2023-09-11
Revised date: 2023-11-01
Online published: 2026-03-11
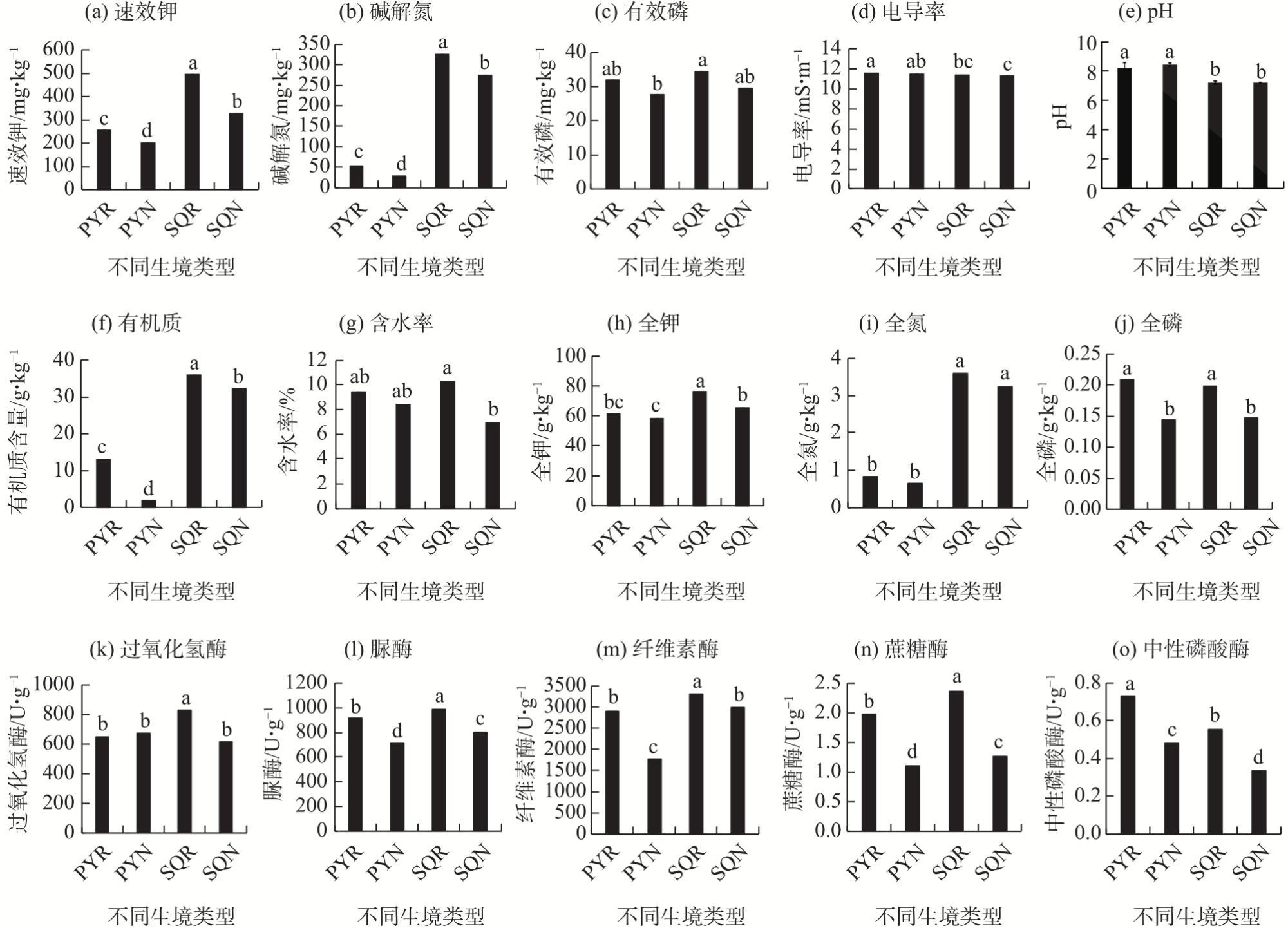
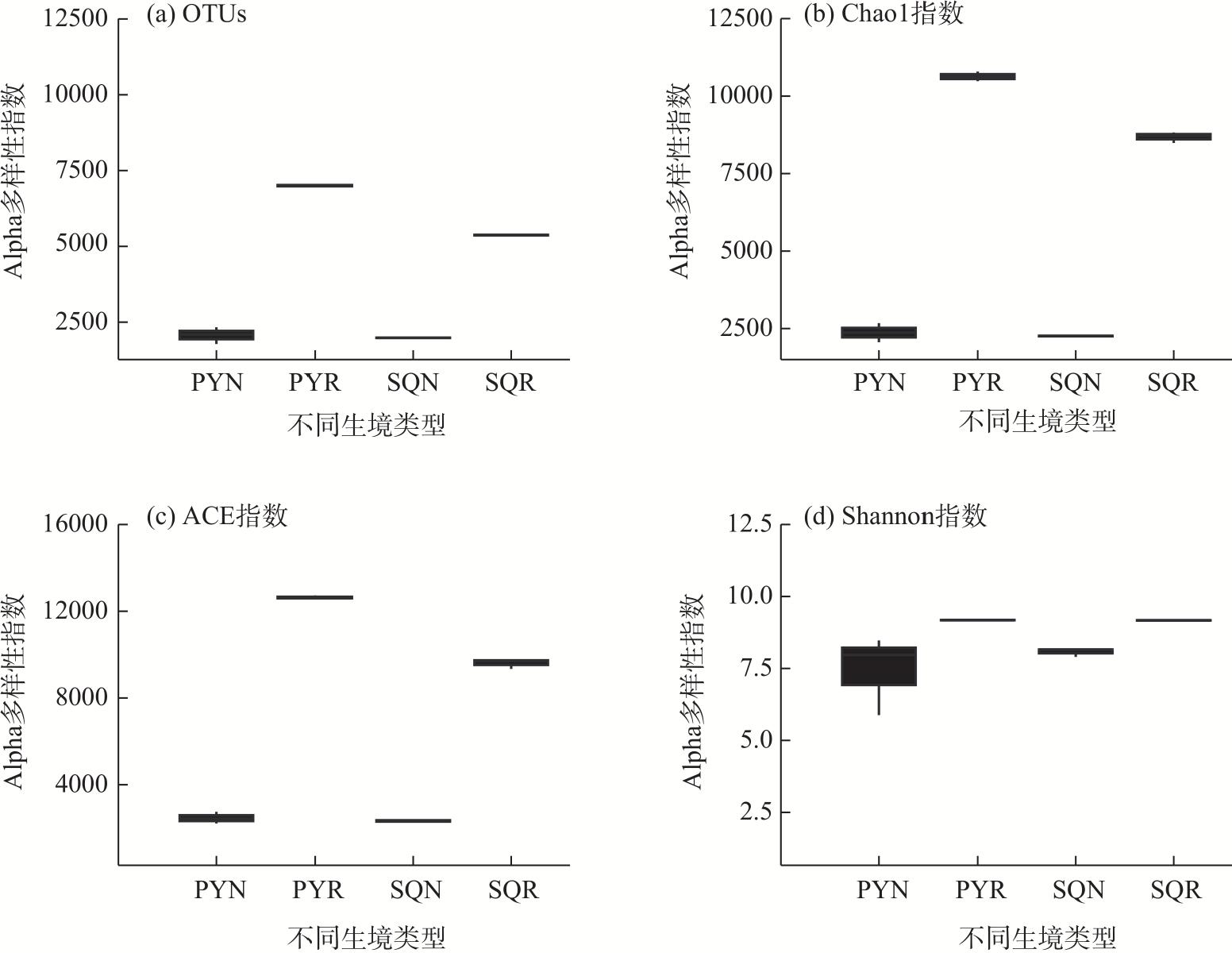
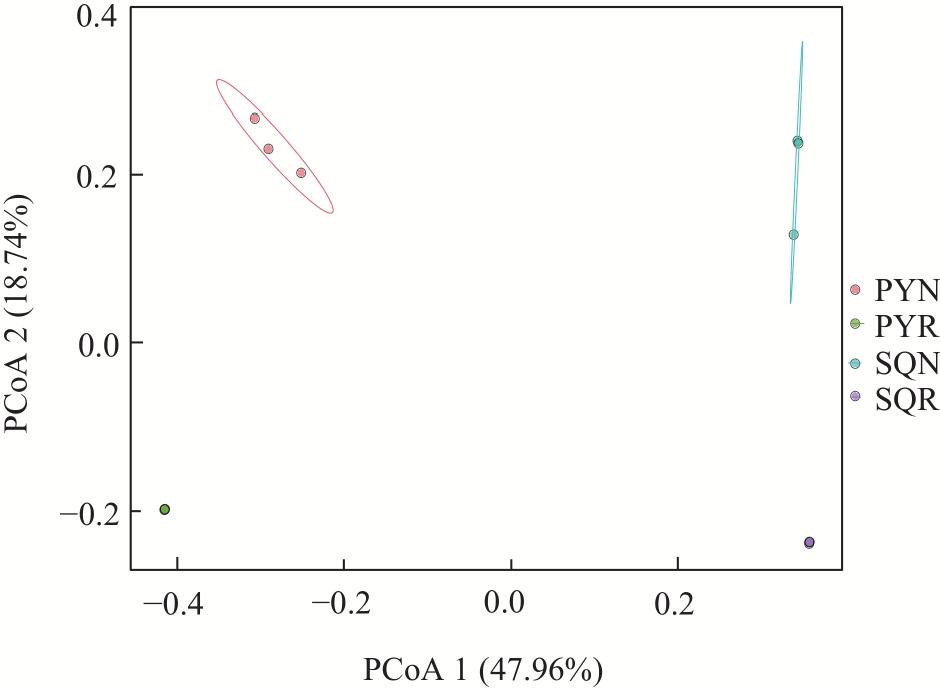
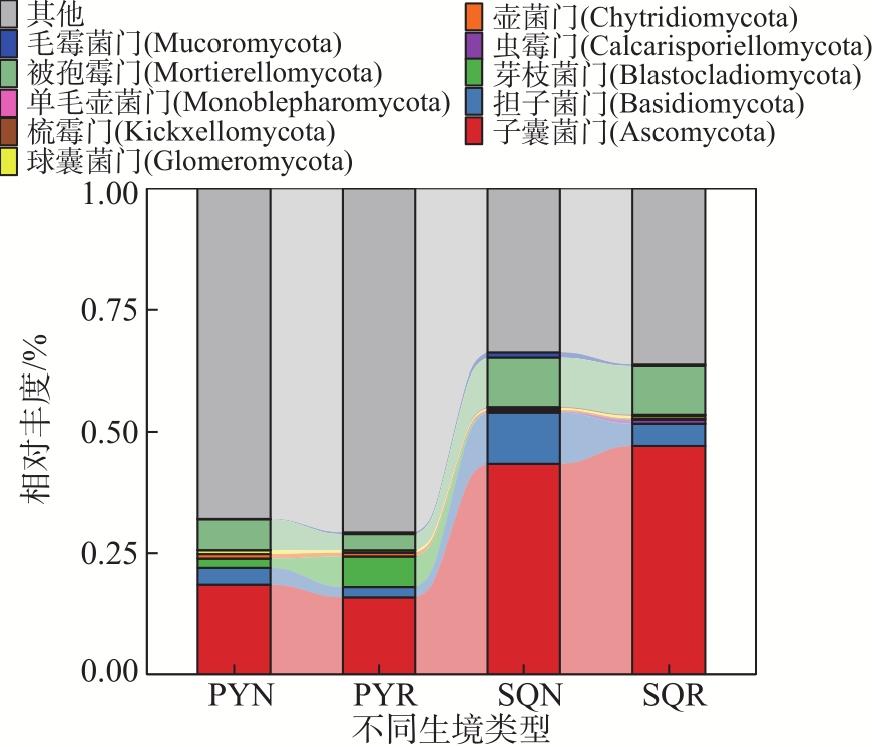
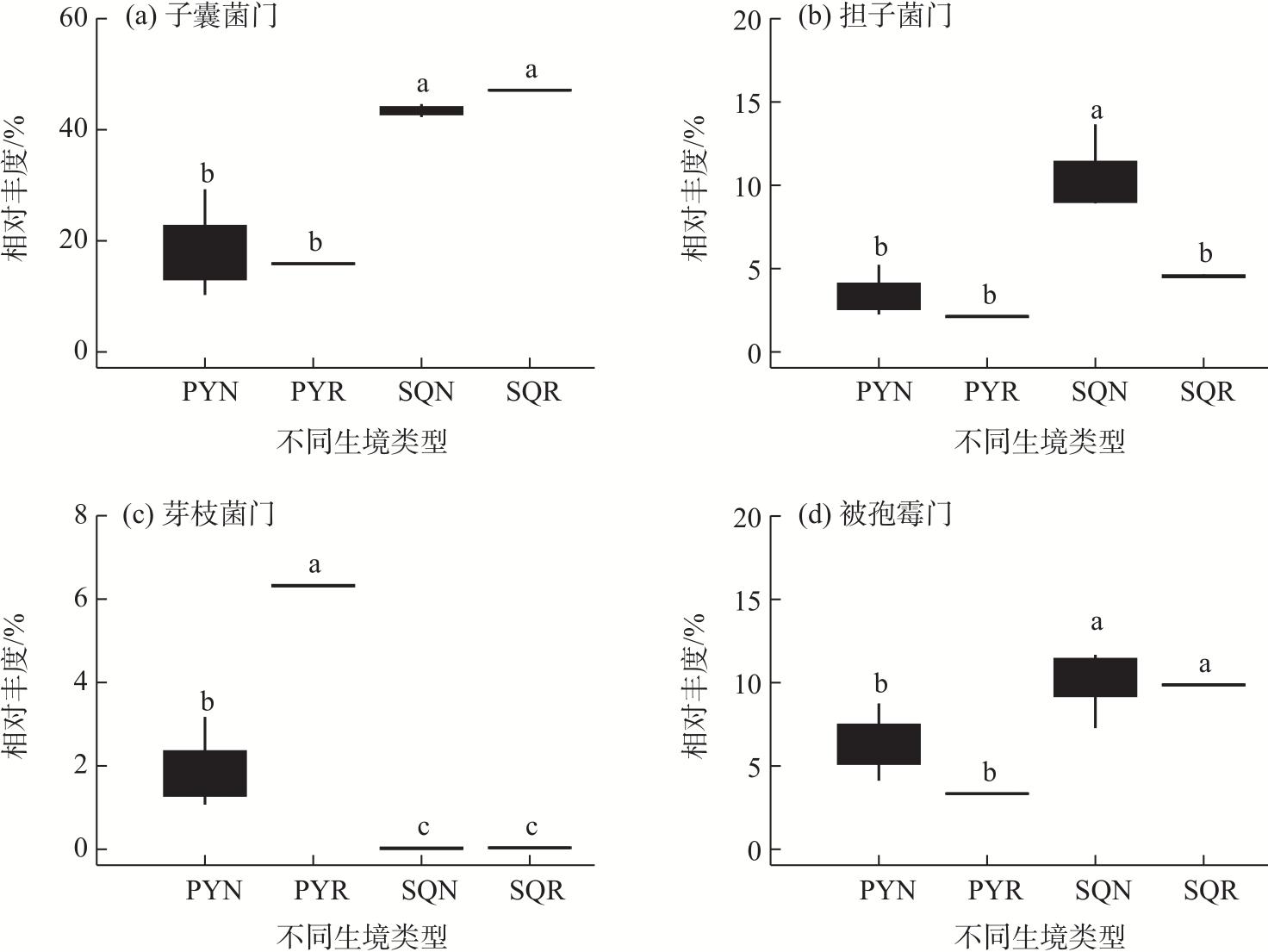
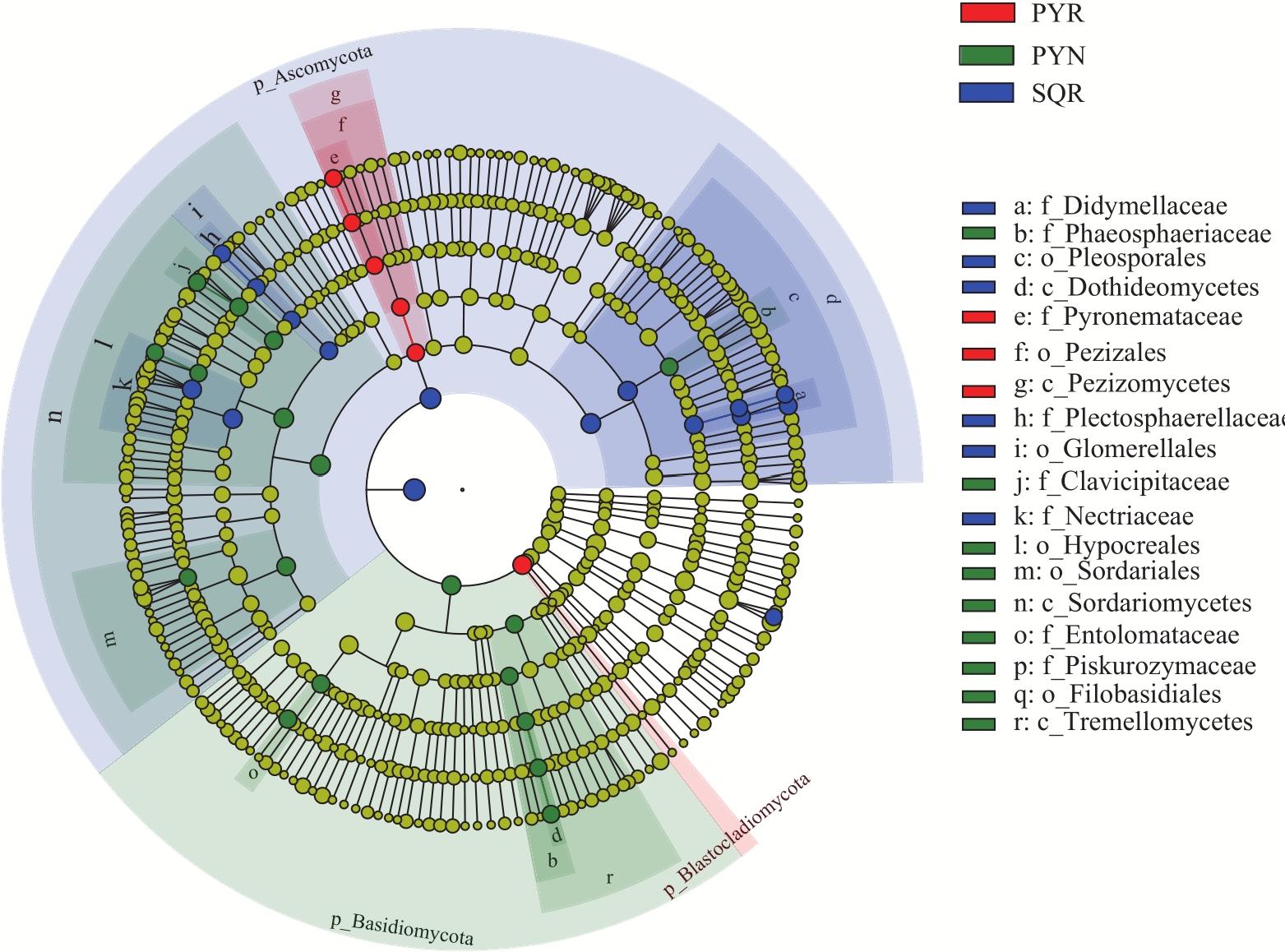
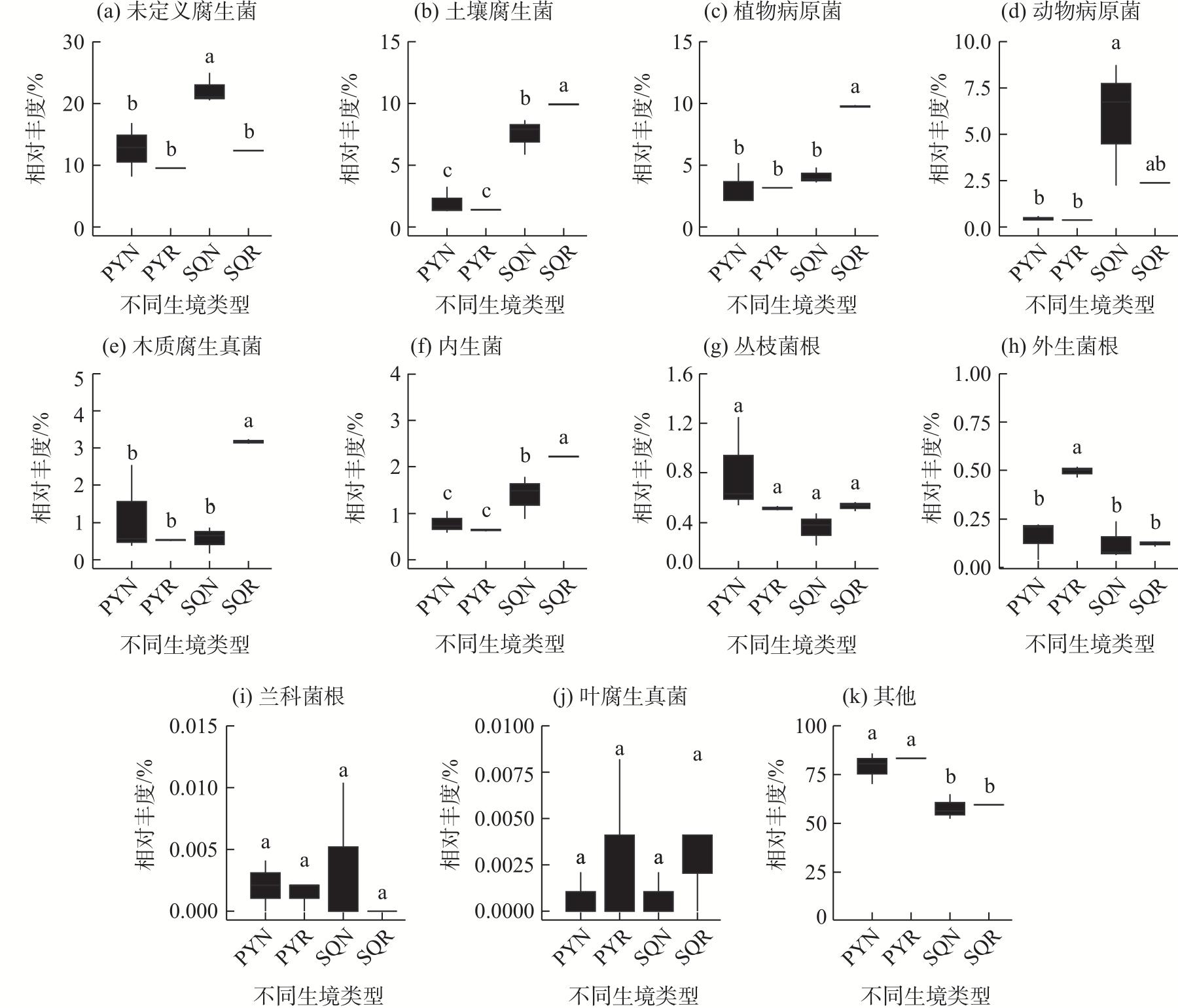
In agricultural ecosystems, rhizosphere microbial communities serve as the driving force for the successful cycling and transformation of nutrients and organic matter in soil, exerting a significant impact on the growth, health, and nutritional status of crops. To investigate the diversity of rhizosphere fungi in different habitats in the northern Tianshan Mountains and the driving factors behind their variations, this study employed high-throughput sequencing to analyze the diversity, structures, functions, and driving factors of rhizosphere and non-rhizosphere fungi in alfalfa rhizosphere soils in mountainous and plain habitats. The results revealed that: (1) Soil nitrogen, potassium, organic matter, and enzyme activity in the mountainous alfalfa rhizosphere soil were significantly higher than those in plain alfalfa rhizosphere soil, while electrical conductivity and pH were significantly lower than those in plain alfalfa rhizosphere soil. (2) Significant differences were observed in the fungal communities in mountainous and plain soils, with the operational taxonomic units (OTUs), Chao1, and ACE indices of plain alfalfa rhizosphere soil being significantly higher than those of mountainous alfalfa rhizosphere soil; additionally, the OTUs, Chao1, ACE, and Shannon indices showed significantly higher values in rhizosphere soils than those in non-rhizosphere soils. (3) The dominant fungal phyla in the region were Ascomycota, Mortierellomycota, Basidiomycota, and Blastocladiomycota, with notable differences in their distribution across different habitats. (4) Based on FUNGuild’s fungal functional predictions, pathogenic fungi in mountainous areas were significantly higher than those in plain areas, while arbuscular mycorrhizal fungi were significantly lower in mountainous areas. (5) Redundancy analysis and Mantel tests indicated that soil pH, total nitrogen, total potassium and available potassium were the main driving factors for the differential distribution of fungal community structures in soils.
Man LI , Qizhang DENG , Yuting JING , Yong WU , Chengming ZHANG , Shan XIA , Yaping ZHAI , Pan YANG , Hongling LIU , Li ZHUANG . Distribution characteristics and driving factors of soil fungi between alfalfa roots in different habitats in northern Tianshan Mountains[J]. Arid Land Geography, 2024 , 47(7) : 1187 -1198 . DOI: 10.12118/j.issn.1000-6060.2023.499
图2 不同生境苜蓿根际土壤真菌多样性注:OTUs为操作分类单元,代表一组相似序列的聚类;Chao1是由Chao提出的用Chao1算法估计样本中所含OTU数目的指数;ACE是用来估计群落中含有OTU数目的指数;Shannon指数为香农指数。 Fig. 2 Fungal diversity in rhizosphere soil of alfalfa in different habitats |
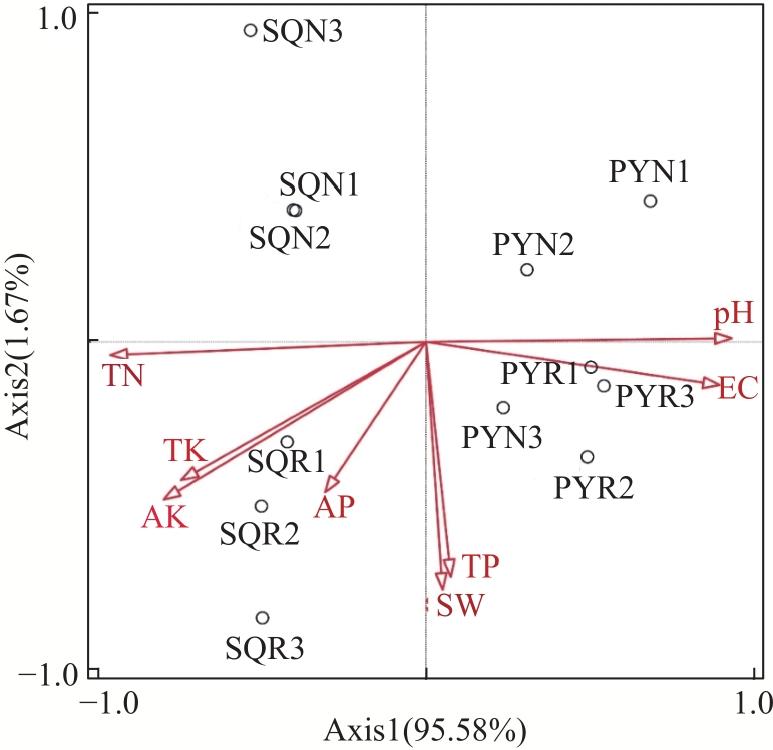
图7 生态环境因子与真菌群落的冗余分析(RDA)Fig. 7 RDA analysis of ecological factors and fungal communities |
表1 蒙特卡洛置换检验Tab. 1 Monte Carlo permutation tests |
| 理化因子 | 重要性排名 | 解释率/% | F值 | P值 |
|---|---|---|---|---|
| 全氮 | 1 | 88.4 | 75.9 | 0.001 |
| pH | 2 | 83.2 | 49.4 | 0.002 |
| 电导率 | 3 | 76.7 | 33.0 | 0.001 |
| 速效钾 | 4 | 61.4 | 15.9 | 0.004 |
| 全钾 | 5 | 53.5 | 11.5 | 0.008 |
| 速效磷 | 6 | 9.5 | 1.0 | 0.296 |
| 全磷 | 7 | 1.5 | 0.2 | 0.733 |
| 含水率 | 8 | 1.2 | 0.1 | 0.781 |
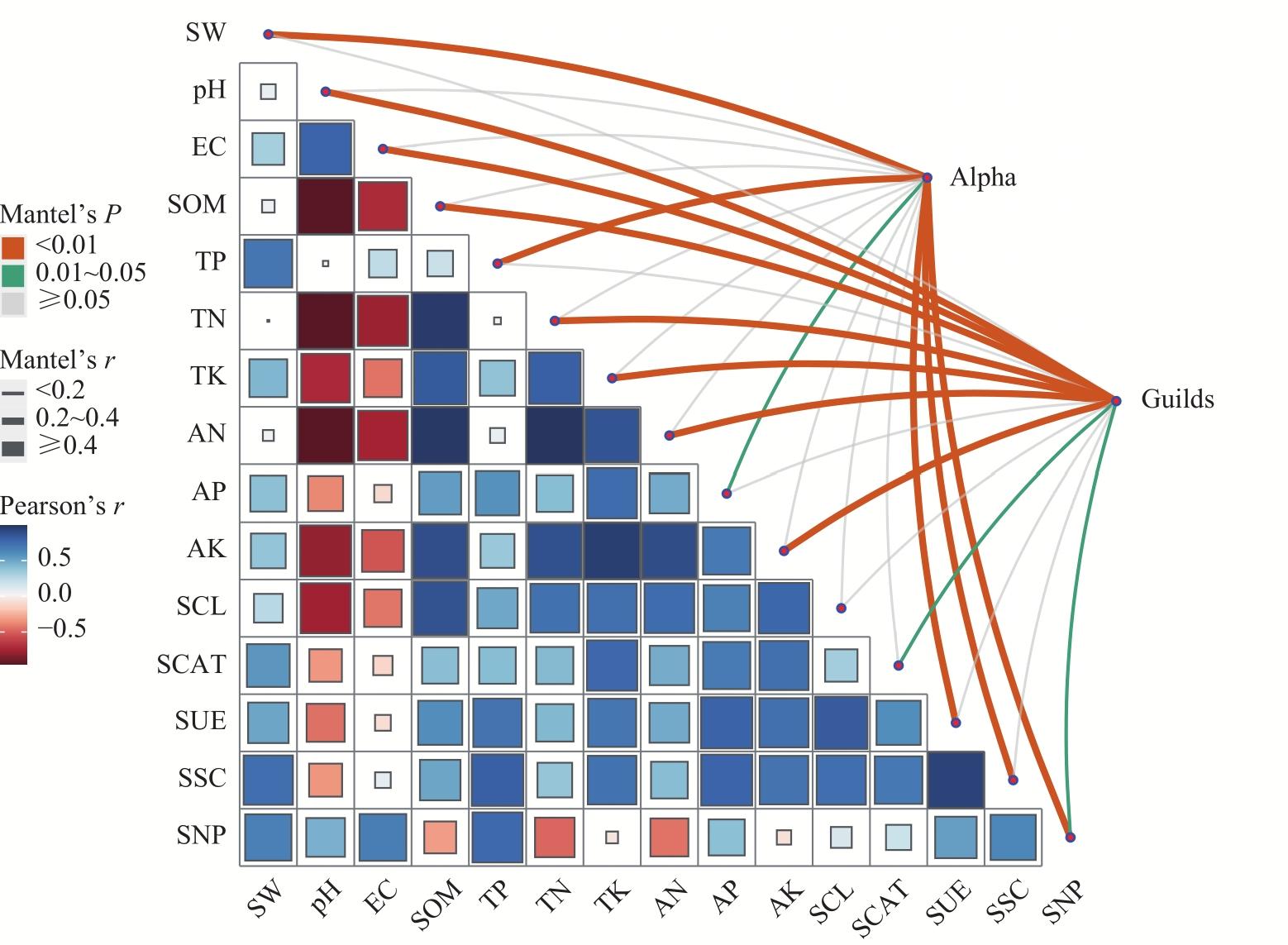
图8 土壤理化环境因子和土壤酶活性与真菌群落Alpha多样性和功能群落之间的Mantel分析注:TK为全钾;TN为全氮;TP为全磷;SOM为有机质;AK为速效钾;AN为碱解氮;AP为有效磷;SW为土壤含水率;EC为电导率;SCAT为过氧化氢酶;SUE为脲酶;SCL为纤维素酶;SSC为蔗糖酶;SNP为土壤中性磷酸酶。Mantel’s P是一种检验2个矩阵之间的相关性是否显著的方法,其值表示相关性检验的显著性水平;Mantel’s r是Mantel 检验的相关系数,用于衡量2个距离矩阵之间的相关性程度;Pearson’s r是皮尔逊相关系数,用于衡量2个连续变量之间的线性关系强度和方向。 Fig. 8 Mantel analysis of the relationship between soil physicochemical environmental factors, soil enzyme activity, and fungal community Alpha diversity and functional communities |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
周霞, 魏杨, 李东嵘, 等. 黄土区紫花苜蓿根系对土体抗剪性能的影响[J]. 中国水土保持科学, 2019, 17(2): 53-59.
[
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
瞿宋林, 吴一凡, 刘忠宽, 等. 丛枝菌根真菌对紫花苜蓿生长发育特性的影响[J]. 草地学报, 2022, 30(10): 2529-2534.
[
|
| [12] |
王倩倩, 陆嘉惠, 张洁, 等. 不同分布区胀果甘草原生境土壤微生物群落结构特征及其影响因素[J]. 生态学报, 2022(23): 1-16.
[
|
| [13] |
王晓春, 高婷. 盐碱地紫花苜蓿根际土壤真菌多样性分析[J]. 农业与技术, 2023, 43(24): 9-12.
[
|
| [14] |
翟亚萍, 王绍明, 刘鸯, 等. 不同种植地苜蓿根际土壤细菌群落结构多样性差异分析[J]. 新疆农业科学, 2021, 58(5): 955-964.
[
|
| [15] |
鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000.
[
|
| [16] |
关松荫. 土壤酶及其研究法[M]. 北京: 农业出版社, 1986.
[
|
| [17] |
韩博远, 张闻, 胡芳雨, 等. 模拟及实际根系分泌物对芘污染土壤微生物群落的影响[J]. 环境科学, 2022, 43(2): 1077-1088.
[
|
| [18] |
邓莹莲, 赵长林. 基于高通量测序分析云南大理剑川羊肚菌土壤细菌群落结构及多样性[J]. 中国农学通报, 2023, 39(10): 79-87.
[
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
徐飞, 张拓, 怀宝东, 等. 土地利用变化对松花江下游湿地土壤真菌群落结构及功能的影响[J]. 环境科学, 2021, 42(5): 2531-2540.
[
|
| [24] |
贾丹, 王琪瑶, 李云红, 等. 小兴安岭红松林土壤理化性质及酶活性的研究[J]. . 林业科技, 2023, 48(3): 23-26.
[
|
| [25] |
刘芳, 汪航飞, 蒲春燕, 等. 不同施肥对葡萄苗根际微生物量、土壤酶活性和生理的影响[J]. 四川农业大学学报, 2023, 41(2): 318-324.
[
|
| [26] |
孙慧, 张建锋, 许华森, 等. 余姚滨海不同盐碱度土壤微生物群落组成及土壤酶活性的变化[J]. 应用生态学报, 2016, 27(10): 3361-3370.
[
|
| [27] |
刘子涵, 黄方园, 黎景来, 等. 覆盖模式对旱作农田土壤微生物多样性及群落结构的影响[J]. 生态学报, 2021, 41(7): 2750-2760.
[
|
| [28] |
高彦婷, 张芮, 董博, 等. 垄沟覆盖集雨模式对玉米根际土壤微生物多样性的影响[J]. 干旱区地理, 2024, 47(3): 413-423.
[
|
| [29] |
于天赫, 张乃莉, 于爽, 等. 北京城市公园常见乔木土壤真菌群落特征及影响因素[J]. 生态学报, 2021, 41(5): 1835-1845.
[
|
| [30] |
姜霓雯, 梁辰飞, 张勇, 等. 土地利用变化后不同种植年限香榧土壤微生物群落的组成及多样性[J]. 环境科学, 2021, 43(1): 1-14.
[
|
| [31] |
周煜杰, 贾夏, 赵永华, 等. 秦岭火地塘真菌群落海拔分布格局[J]. 应用生态学报, 2021, 32(7): 2589-2596.
[
|
| [32] |
|
| [33] |
|
| [34] |
王诗慧, 常顺利, 李鑫, 等. 天山林区土壤真菌多样性及其群落结构[J]. 生态学报, 2021, 41(1): 124-134.
[
|
| [35] |
杨盼, 翟亚萍, 赵祥, 等. AM真菌和根瘤菌互作对苜蓿根际土壤真菌群落结构的影响及功能预测[J]. 草业科学, 2020, 37(9): 1669-1680.
[
|
| [36] |
王小玲, 马琨, 伏云珍, 等. 免耕覆盖及有机肥施用对土壤真菌群落组成及多样性的影响[J]. 应用生态学报, 2020, 31(3): 890-898.
[
|
| [37] |
刘震, 徐玉鹏, 王秀领, 等. 黑龙港苜蓿根际土壤真菌群落结构及多样性分析[J]. 江苏农业科学, 2021, 49(10): 197-201.
[
|
| [38] |
孙俊奇, 杨超, 金静, 等. 被孢霉真菌代谢液对苜蓿、马唐、稗草的影响[J]. 中国草地学报, 2023, 45(11): 92-98.
[
|
| [39] |
袁军. 中国西南地区亚隔孢壳科(Didymellaceae)植物病原真菌分子系统学研究[D]. 贵阳: 贵州大学, 2021.
[
|
| [40] |
赵鹏, 罗晶, 庄文颖. COI基因作为丛赤壳科真菌DNA条形码的测试[J]. 菌物学报, 2012, 31(2): 243-250.
[
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
徐丽娟, 刁志凯, 李岩, 等. 菌根真菌的生理生态功能[J]. 应用生态学报, 2012, 23(1): 285-292.
[
|
| [45] |
|
| [46] |
罗正明, 刘晋仙, 暴家兵, 等. 五台山亚高山土壤真菌海拔分布格局与构建机制[J]. 生态学报, 2020, 40(19): 7009-7017.
[
|
| [47] |
刘晓勤, 张锐利. 新疆红枣黑斑病根际土壤真菌群落组成及多样性的分析[J]. 北方园艺, 2020(14): 94-102.
[
|
| [48] |
陈永成, 李肖, 蔡宜东, 等. 昆仑山沙区旱作紫花苜蓿根际/非根际土壤理化性质及真菌群落特征[J]. 草业科学, 2023, 40(5): 1232-1242.
[
|
| [49] |
张晓丽, 张宏媛, 卢闯, 等. 河套灌区不同秋浇年限对土壤细菌群落的影响[J]. 中国农业科学, 2019, 52(19): 3380-3392.
[
|
| [50] |
|
| [51] |
刘震, 徐玉鹏, 黄伟, 等. 苜蓿连作对盐碱土壤微生物群落结构的影响[J]. 作物研究, 2020, 34(6): 557-562.
[
|
| [52] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}