
Biomass and soil nutrient characteristics of five dominant plant species in the desert grassland of the northern foothills of the Qilian Mountains
Received date: 2023-06-09
Revised date: 2023-07-26
Online published: 2024-06-20
To clarify the relationship between plant biomass and soil nutrients of dominant plants in desert grasslands, this study selected five dominant plant species: Peganum harmala, Setaria viridis, Festuca sinensis, Puccinellia distans, and Agropyron cristatum. We measured their biomass and root-zone soil nutrients to explore biomass allocation and its relationship with soil nutrients. The results are as follows: (1) There were significant differences in total biomass and root-shoot ratio among the five dominant plant species (P<0.05), with Peganum harmala having the highest total biomass and Puccinellia distans having the lowest. The perennial plants Peganum harmala, Puccinellia distans, Festuca sinensis, and Agropyron cristatum had most of their biomass allocated below ground, whereas the annual plant Setaria viridis had most of its biomass above ground. The order of root-shoot ratio among the five plants was as follows: Puccinellia distans>Agropyron cristatum>Festuca sinensis>Peganum harmala>Setaria viridis. (2) There were significant differences (P<0.05) in root-zone soil organic carbon, available nitrogen, available potassium, total nitrogen, total phosphorus, total potassium, and their stoichiometric characteristics among the five plant species. The order of soil C:N ratio among the five plants was as follows: Puccinellia distans>Setaria viridis>Agropyron cristatum>Peganum harmala>Festuca sinensis. (3) Variations in plant biomass, root-shoot ratio, and soil nutrients varied among the plants. The root-zone soil total potassium of Peganum harmala, Setaria viridis, and Festuca sinensis and the root-zone soil moisture of Puccinellia distans and Agropyron cristatum exhibited weak variation, whereas the other plant characteristics, soil nutrients, and stoichiometric characteristics exhibited moderate variation. The biomass of the five dominant plant species exhibited a positive correlation with the root-zone soil available nitrogen and total potassium (P<0.05). The allocation of plant biomass and soil nutrient composition significantly vary among different species and life histories in the desert grassland ecosystem. In the future, it will be necessary to restore degraded desert ecosystems by applying appropriate fertilization based on the nutrient requirements of different dominant plant species.
Zhiming ZHANG , Xiaomei SUN , Duanhong BAO , Baohui YAO , Zhicheng WANG , Junhu SU . Biomass and soil nutrient characteristics of five dominant plant species in the desert grassland of the northern foothills of the Qilian Mountains[J]. Arid Land Geography, 2024 , 47(4) : 662 -671 . DOI: 10.12118/j.issn.1000-6060.2023.272
表1 祁连山北麓5种植物生物量及根冠比Tab. 1 Biomass and root-to-shoot ratio of five species at the northern foothills of the Qilian Mountains |
| 物种 | 样本数 | 地上生物量 | 地下生物量 | 总生物量 | 根冠比 |
|---|---|---|---|---|---|
| 骆驼蓬(Peganum harmala) | 25 | 13.57a | 21.30a | 34.88a | 1.57cd |
| 狗尾草(Setaria viridis) | 25 | 0.59b | 0.52b | 1.10b | 0.92d |
| 中华羊茅(Festuca sinensis) | 25 | 0.26b | 1.18b | 1.44b | 2.26c |
| 碱茅(Puccinellia distans) | 25 | 0.07b | 0.24b | 0.32b | 5.19a |
| 冰草(Agropyron cristatum) | 25 | 0.42b | 1.12b | 1.55b | 3.29b |
注:不同小写字母表示差异显著(P<0.05)。下同。 |
表2 祁连山北麓5种植物根际土壤养分及其化学计量特征Tab. 2 Rhizosphere soil nutrients and their stoichiometric characteristics of five plants at the northern foothills of the Qilian Mountains |
| 土壤指标 | 狗尾草 | 中华羊茅 | 碱茅 | 冰草 | 骆驼蓬 |
|---|---|---|---|---|---|
| SWC/% | 16.2±1.05a | 10.4±1.68a | 13.36±0.46a | 16.45±0.81a | 14.84±3.95a |
| AP/mg·kg-1 | 10.82±0.44a | 11.41±6.46a | 12.34±0.24a | 11.35±0.83a | 10.55±0.29a |
| AK/mg·kg-1 | 3.54±0.41b | 1.07±0.51c | 1.58±2.31c | 7.13±1.70a | 2.85±4.50b |
| AN/mg·kg-1 | 53.99±21.38c | 95.64±10.68b | 97.31±31.87b | 98.3±0.59b | 137.27±14.58a |
| TN/g·kg-1 | 0.29±0.17b | 0.51±0.15a | 0.24±0.10b | 0.28±0.12b | 0.31±0.07b |
| TP/g·kg-1 | 2.30±0.44a | 2.59±0.13a | 0.75±0.13b | 0.84±0.27b | 0.73±0.04b |
| TK/g·kg-1 | 10.38±0.56a | 6.21±0.62b | 5.92±0.63b | 6.15±0.71b | 10.03±0.92a |
| OC/g·kg-1 | 49.93±9.44ab | 26.33±7.89c | 59.59±12.45a | 37.79±15.13bc | 22.26±10.72c |
| N:P | 0.13±0.06b | 0.20±0.07b | 0.32±0.14ab | 0.33±0.20a | 0.42±0.08a |
| C:K | 2.79±0.61bc | 2.46±0.99bc | 5.84±1.16a | 3.56±1.60b | 1.29±0.75c |
| C:P | 12.59±1.00bc | 5.90±1.80c | 45.81±13.41a | 26.19±2.56bc | 17.59±7.63c |
| C:N | 99.34±39.05b | 30.24±28.25b | 145.07±55.48a | 78.72±34.49b | 42.00±13.62b |
注:SWC为土壤含水量;AP为速效磷;AK为速效钾;AN为碱解氮;TN为全氮;TP为全磷;TK为全钾;OC为有机碳;C:N为碳氮比;C:P为碳磷比;N:P为氮磷比;C:K为碳钾比。下同。 |
表3 祁连山北麓5种植物根际土壤养分、生物量 及根冠比变异系数Tab. 3 Coefficients of variation of soil nutrients, biomass and root-shoot ratio of five plants at the northern foothills of the Qilian Mountains |
| 土壤指标 | 变异系数 | ||||
|---|---|---|---|---|---|
| 骆驼蓬 | 狗尾草 | 中华羊茅 | 碱茅 | 冰草 | |
| OC | 48.13 | 18.91 | 29.98 | 20.88 | 40.03 |
| TN | 24.10 | 44.50 | 35.15 | 47.80 | 33.74 |
| TP | 5.64 | 19.18 | 5.10 | 16.62 | 32.20 |
| TK | 9.17 | 5.42 | 9.96 | 10.65 | 11.51 |
| C:N | 33.09 | 46.91 | 68.72 | 29.86 | 50.06 |
| C:P | 43.80 | 7.94 | 30.50 | 28.51 | 9.92 |
| N:P | 18.32 | 32.94 | 38.14 | 49.34 | 44.80 |
| C:K | 56.58 | 21.79 | 39.54 | 19.73 | 44.05 |
| AN | 11.16 | 35.25 | 11.17 | 32.75 | 0.60 |
| AP | 42.62 | 3.83 | 4.47 | 18.71 | 14.96 |
| AK | 10.19 | 12.49 | 137.04 | 15.40 | 11.61 |
| SWC | 26.59 | 6.48 | 16.14 | 3.44 | 4.95 |
| Ab | 8.11 | 32.18 | 34.19 | 67.27 | 18.35 |
| Bb | 13.97 | 47.50 | 97.78 | 12.56 | 66.12 |
| Tb | 11.62 | 30.39 | 85.48 | 20.55 | 51.97 |
| R:S | 6.20 | 42.80 | 80.19 | 46.90 | 53.52 |
注:Ab为地上生物量;Bb为地下生物量;Tb为总生物量;R:S为根冠比。下同。 |
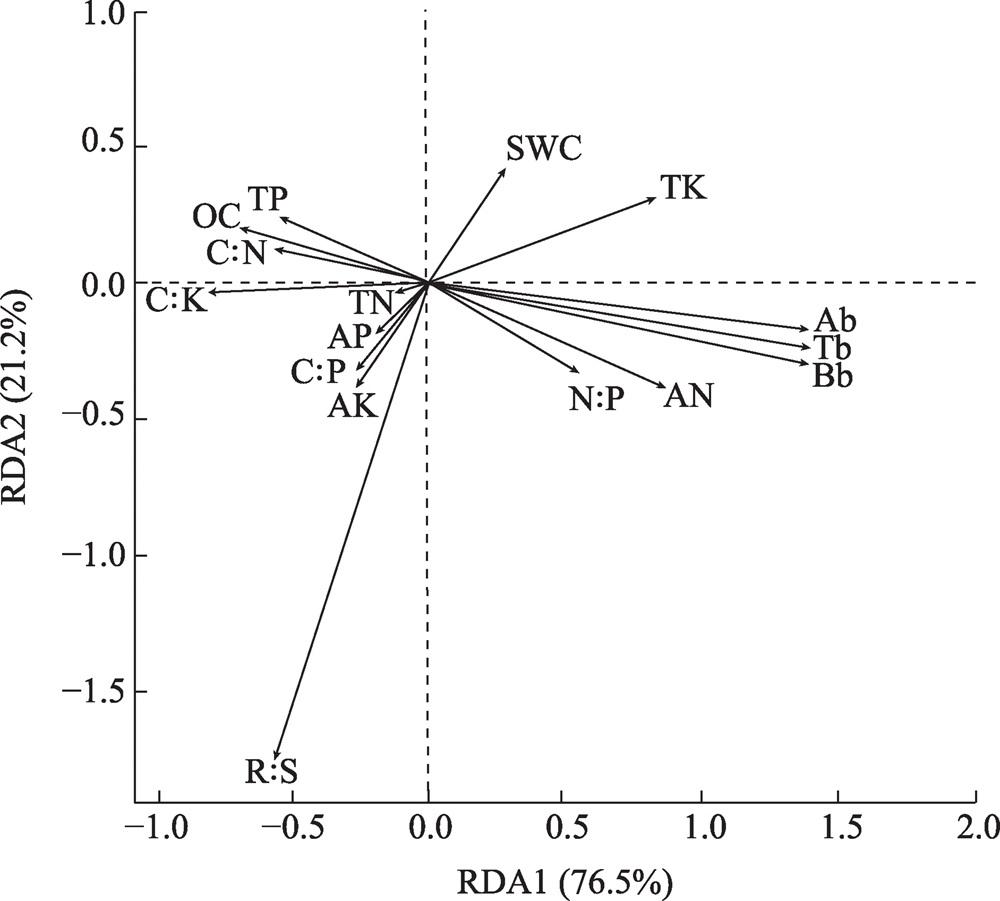
图2 植物生物量和土壤养分特征的RDA排序图注:RDA1、RDA2分别为冗余分析第一排序轴和第二排序轴。 Fig. 2 RDA ranking plot of plant biomass and soil nutrient characteristics |
表4 土壤因子与植物生物量RDA结果Tab. 4 RDA results of soil factors and plant biomass |
| 内容 | RDA1 | RDA2 |
|---|---|---|
| 特征值 | 3.0589 | 0.8496 |
| 可解释变量累计百分比 | 0.7647 | 0.2124 |
| 可解释拟合变量(累计) | 0.7647 | 0.9771 |
| 蒙特卡罗置换检验 | P<0.05 | |
注:RDA1、RDA2分别为冗余分析第一排序轴和第二排序轴。下同。 |
表5 土壤因子和植物生物量RDA排序相关系数Tab. 5 RDA ranking correlation coefficient between soil factors and plant biomass |
| 土壤因子 | RDA1 | RDA2 | R2 | Pr |
|---|---|---|---|---|
| SWC | 0.47101 | 0.88213 | 0.1952 | 0.245 |
| AK | -0.47991 | -0.87732 | 0.1677 | 0.327 |
| AP | -0.62465 | -0.78091 | 0.0503 | 0.726 |
| AN | 0.87023 | -0.49264 | 0.4884 | 0.022 |
| TN | -0.92070 | -0.39028 | 0.0089 | 0.951 |
| TP | -0.87356 | 0.48671 | 0.1915 | 0.280 |
| TK | 0.89323 | 0.44961 | 0.4301 | 0.046 |
| OC | -0.94157 | 0.33682 | 0.2649 | 0.156 |
| N:P | 0.78968 | -0.61352 | 0.2386 | 0.193 |
| C:K | -0.99676 | -0.08045 | 0.3200 | 0.095 |
| C:P | -0.54229 | -0.84019 | 0.1281 | 0.411 |
| C:N | -0.95473 | 0.29746 | 0.1397 | 0.429 |
注:R2为决定系数;Pr为相关性的显著性检验。 |
| [1] |
沈海花, 朱言坤, 赵霞, 等. 中国草地资源的现状分析[J]. 科学通报, 2016, 61(2): 139-154.
[
|
| [2] |
|
| [3] |
于钊, 李奇铮, 王培源, 等. 退化和恢复过程驱动的荒漠草地生态系统有机碳密度变化[J]. 中国沙漠, 2022, 42(2): 215-222.
[
|
| [4] |
|
| [5] |
|
| [6] |
哈斯, 盖志毅. 基于政策过程的我国荒漠化治理研究述评[J]. 科学管理研究, 2021, 39(2): 29-36.
[
|
| [7] |
武海霞, 王则宇, 尹强, 等. 我国北方荒漠草原退化研究进展[J]. 内蒙古林业科技, 2017, 43(2): 58-62.
[
|
| [8] |
张继辉, 蔡道雄, 卢立华, 等. 不同林龄柚木人工林土壤生态化学计量特征[J]. 生态学报, 2020, 40(16): 5718-5728.
[
|
| [9] |
张军, 安邦. 安徽省潜山市土壤养分地球化学特征及其影响因素[J]. 能源技术与管理, 2022, 47(4): 190-194.
[
|
| [10] |
李巧玲, 阎欣, 吴秀芝, 等. 荒漠草原沙漠化对土壤无机碳和有机碳的影响[J]. 水土保持学报, 2019, 33(1): 98-103, 110.
[
|
| [11] |
唐庄生, 安慧, 邓蕾, 等. 荒漠草原沙漠化植物群落及土壤物理变化[J]. 生态学报, 2016, 36(4): 991-1000.
[
|
| [12] |
唐庄生, 安慧, 上官周平. 荒漠草原沙漠化对土壤养分与植被根冠比的影响[J]. 草地学报, 2015, 23(3): 463-468.
[
|
| [13] |
赵雯, 黄来明. 高寒山区不同土地利用类型土壤养分化学计量特征及影响因素[J]. 生态学报, 2022, 42(11): 4415-4427.
[
|
| [14] |
刘梦云, 吴健利, 刘丽雯, 等. 黄土台塬土地利用方式对土壤水稳性团聚体稳定性影响[J]. 自然资源学报, 2016, 31(9): 1564-1576.
[
|
| [15] |
莫保儒, 王子婷, 蔡国军, 等. 半干旱黄土区成熟柠条林地剖面土壤水分环境及影响因子研究[J]. 干旱区地理, 2014, 37(6): 1207-1215.
[
|
| [16] |
王春燕, 燕霞, 顾梦鹤. 黄土高原弃耕地植被演替及其对土壤养分动态的影响[J]. 草业学报, 2018, 27(11): 26-35.
[
|
| [17] |
王向涛, 张世虎, 陈懂懂, 等. 不同放牧强度下高寒草甸植被特征和土壤养分变化研究[J]. 草地学报, 2010, 18(4): 510-516.
[
|
| [18] |
王占义, 赵向玲, 王成杰, 等. 不同牧马强度对荒漠草原植物群落特征的影响[J]. 草原与草业, 2023, 35(1): 10-16.
[
|
| [19] |
张馨文, 安慧, 杜忠毓, 等. 氮添加对宁夏荒漠草原植物初级生产力的影响机制[J]. 生态学报, 2023, 43(6): 2476-2487.
[
|
| [20] |
图纳热, 红梅, 闫瑾, 等. 降水变化和氮沉降对荒漠草原土壤细菌群落结构及酶活性的影响[J]. 农业环境科学学报, 2023, 42(2): 403-413.
[
|
| [21] |
付爱良, 杨刚, 郑晓红, 等. 新疆荒漠草地退化的原因分析与修复对策探讨[J]. 草食家畜, 2009(1): 6-7.
[
|
| [22] |
阿克塞县地方史志办公室. 阿克塞年鉴[M]. 兰州: 甘肃民族出版社, 2020.
[ Aksay County Local Historical Records Office. Aksay yearbook[M]. Lanzhou: Gansu Ethnic Publishing House, 2020. ]
|
| [23] |
|
| [24] |
鲍士旦. 土壤农业分析[M]. 第3版. 北京: 中国农业出版社, 1999.
[
|
| [25] |
姚宝辉, 王缠, 张倩, 等. 甘南高寒草甸退化过程中土壤理化性质和微生物数量动态变化[J]. 水土保持学报, 2019, 33(3): 138-145.
[
|
| [26] |
景明慧, 贾晓彤, 张运龙, 等. 长期氮添加对内蒙古典型草原植物地上、地下生物量及根冠比的影响[J]. 生态学杂志, 2020, 39(10): 3185-3193.
[
|
| [27] |
崔向慧, 卢琦. 中国荒漠化防治标准化发展现状与展望[J]. 干旱区研究, 2012, 29(5): 913-919.
[
|
| [28] |
杨婷婷, 高永, 吴新宏, 等. 小针茅草原植被地下与地上生物量季节动态及根冠比变化规律[J]. 干旱区研究, 2013, 30(1): 109-114.
[
|
| [29] |
毛伟, 李玉霖, 崔夺, 等. 沙质草地不同生活史植物的生物量分配对氮素和水分添加的响应[J]. 植物生态学报, 2014, 38(2): 125-133.
[
|
| [30] |
姚喜喜, 宫旭胤, 白滨, 等. 祁连山高寒牧区不同类型草地植被特征与土壤养分及其相关性研究[J]. 草地学报, 2018, 26(2): 371-379.
[
|
| [31] |
蔡楠, 孙旭, 刘永宏. 包头黄河湿地不同植物群落土壤养分分布特征[J]. 北方园艺, 2021(10): 80-87.
[
|
| [32] |
朱秋莲, 邢肖毅, 张宏, 等. 黄土丘陵沟壑区不同植被区土壤生态化学计量特征[J]. 生态学报, 2013, 33(15): 4674-4682.
[
|
| [33] |
邵文山, 李国旗, 陈科元, 等. 荒漠草原四种常见植物群落土壤养分及土壤微生物特征[J]. 北方园艺, 2016(15): 161-166.
[
|
| [34] |
王永槐. 退化草地不同功能群生物量与土壤养分的相关性分析[J]. 安徽农业科学, 2014, 42(13): 3975-3977.
[
|
| [35] |
米佳, 刘花, 石晶, 等. 典型草原4种植物根际水分和土壤养分特征研究[J]. 山西大学学报(自然科学版), 2022, 45(4): 1128-1137.
[
|
| [36] |
黄昌勇. 土壤学[M]. 北京: 中国农业出版社, 2000.
[
|
| [37] |
|
| [38] |
|
| [39] |
曹娟, 闫文德, 项文化, 等. 湖南会同3个林龄杉木人工林土壤碳、氮、磷化学计量特征[J]. 林业科学, 2015, 51(7): 1-8.
[
|
| [40] |
郭鑫, 魏天兴, 陈宇轩, 等. 黄土丘陵区典型退耕恢复植被土壤生态化学计量特征[J]. 干旱区地理, 2022, 45(6): 1899-1907.
[
|
| [41] |
汝海丽, 张海东, 焦峰, 等. 黄土丘陵区微地形梯度下草地群落植物与土壤碳、氮、磷化学计量学特征[J]. 自然资源学报, 2016, 31(10): 1752-1763.
[
|
| [42] |
李凯辉, 胡玉昆, 王鑫, 等. 不同海拔梯度高寒草地地上生物量与环境因子关系[J]. 应用生态学报, 2007, 18(9): 2019-2024.
[
|
| [43] |
|
| [44] |
王政权, 张彦东, 王庆成. 水曲柳幼苗根系对土壤养分和水分空间异质性的反应[J]. 植物研究, 1999(3): 329-334.
[
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
潘庆民, 薛建国, 陶金, 等. 中国北方草原退化现状与恢复技术[J]. 科学通报, 2018, 63(17): 1642-1650.
[
|
| [49] |
李永康, 许冬梅, 撒春宁, 等. 乡土物种对退化荒漠草原的改良效果研究[J]. 中国草地学报, 2022, 44(6): 36-45.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}