
Remote Sensing for Natural Resources >
Spatiotemporal trajectory of Spartina alterniflora expansion and its impact on landscape patterns in the Yancheng coastal wetland since the 1980s
Received date: 2023-10-31
Revised date: 2024-06-17
Online published: 2026-06-03
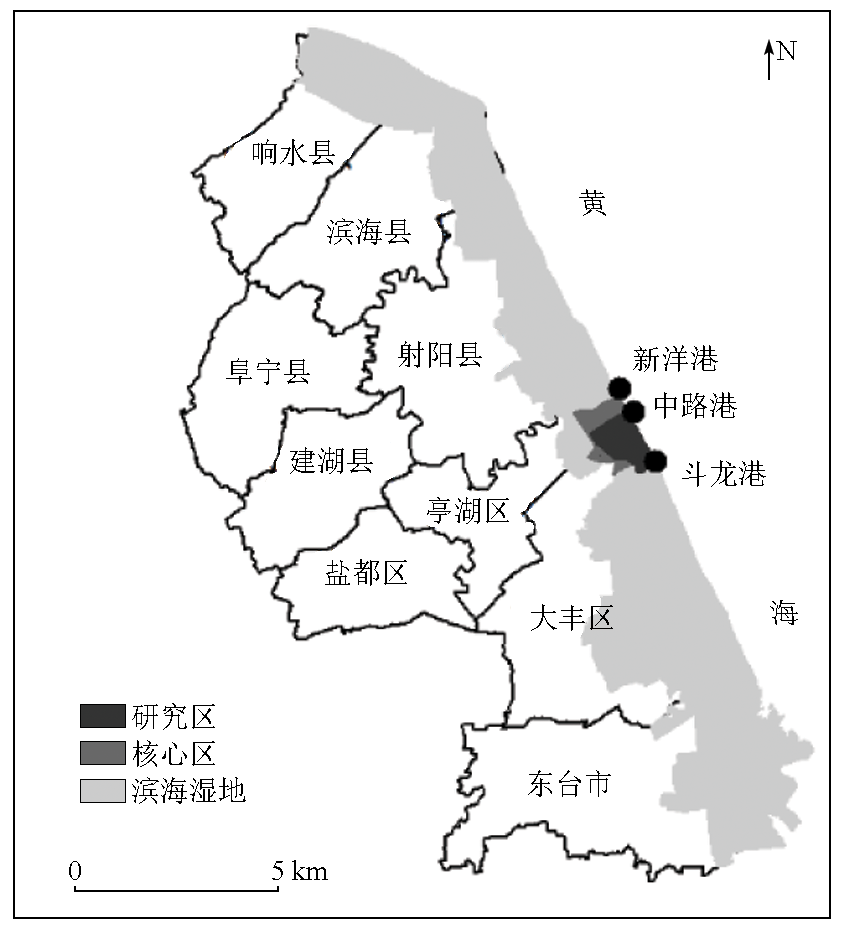
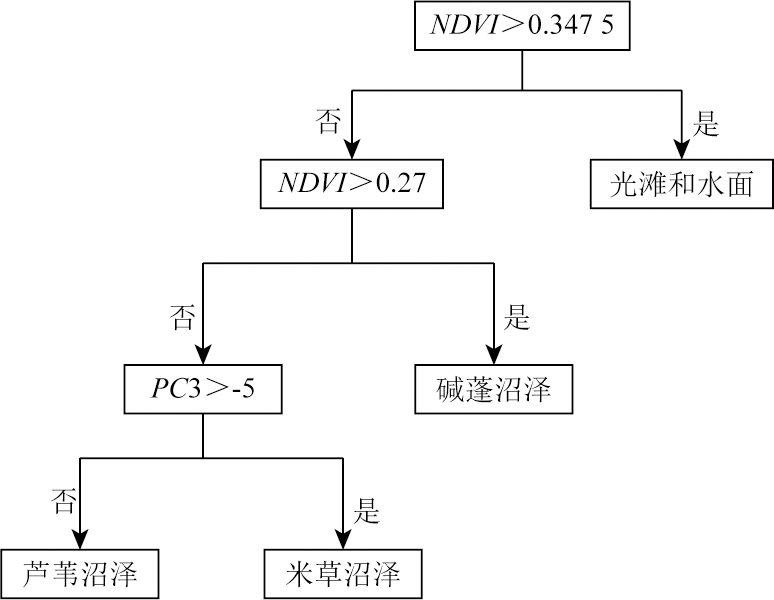
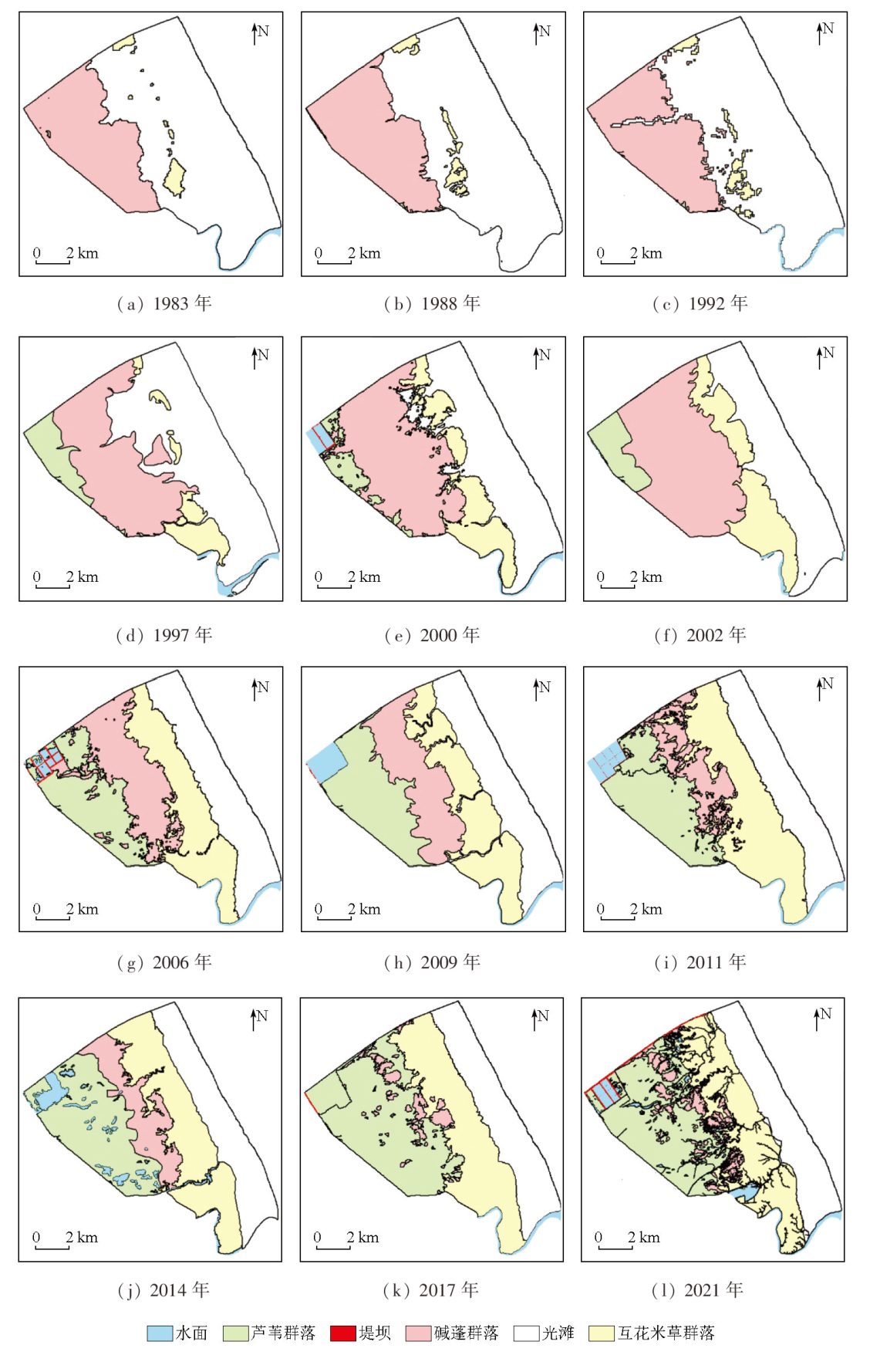
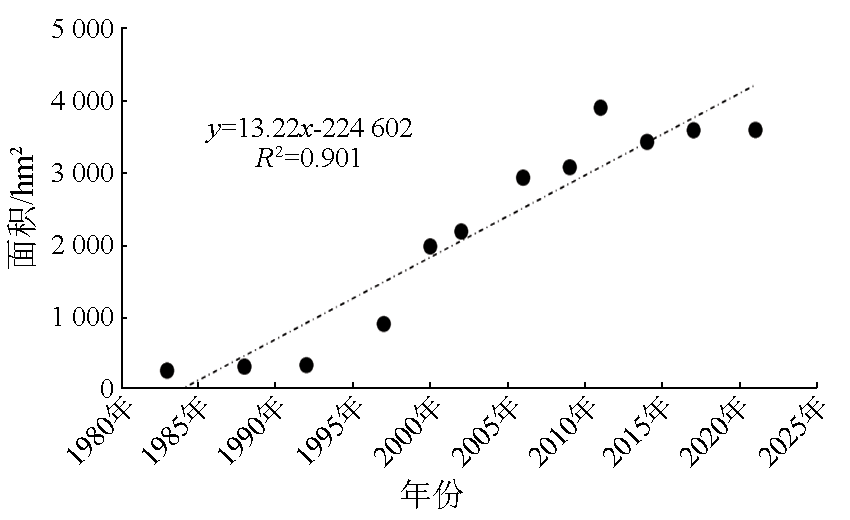
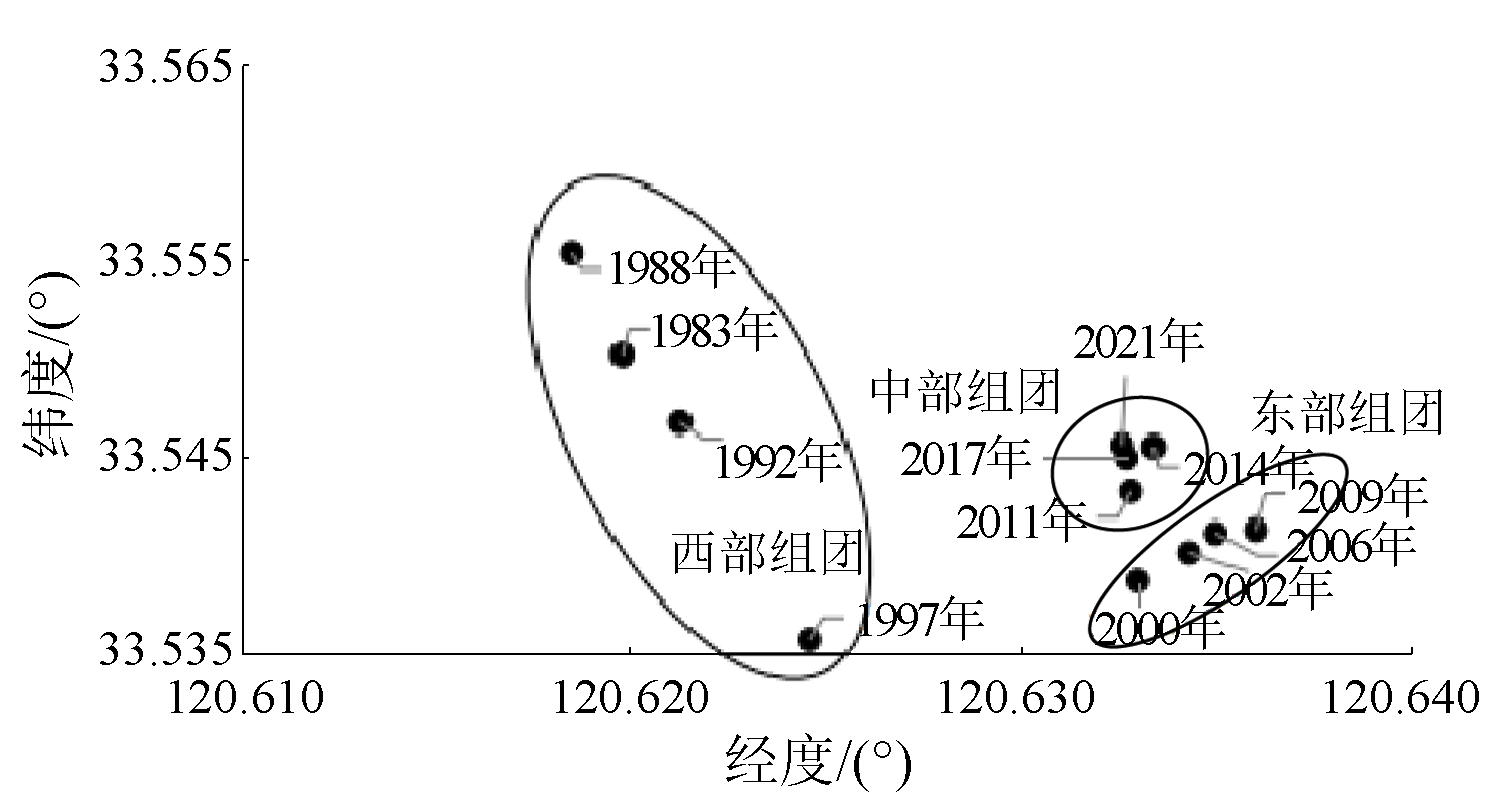
This study investigated the core area of the Jiangsu Yancheng Wetland National Reserve of Rare Birds based on 12 remote sensing images from 1983 to 2021 as data sources. Specifically, this study explored the spatiotemporal trajectory of Spartina alterniflora expansion and its impact on landscape patterns by combining landscape ecology methods with geographical information system (GIS) technology. The results show that from 1983 to 2021, Spartina alterniflora expanded significantly, leading to a 12.365 fold area increase. It experienced initial expansion, accelerated growth, and stagnation stages, which would be succeeded by the control and elimination stage. Its distribution area manifested a significant linear relationship with time. Its spatiotemporal trajectory was characterized by the migration of the western, eastern, and central clusters. In 1983, 1988, 1992, and 1997, the western clusters migrated primarily toward the southeast. In 2000, 2002, 2006, and 2009, the eastern clusters migrated principally toward the northeast. Both the western and eastern clusters showed a seaward trend. In 2011, 2014, 2017, and 2021, the central clusters displayed a significant landward trend despite disorderly migration. Spartina alterniflora expansion resulted in a cumulative contribution rate of 43.352% to regional landscape structure changes. Its contribution was consistent with its expansion stages, showing a low-high-low pattern. The area of Spartina alterniflora was significantly correlated with the regional landscape pattern index. The landscape pattern of Spartina alterniflora was significantly correlated with the regional landscape pattern, with significant correlations between type-scale indices, including largest patch index (LPI), total edge (TE), edge density (ED), and fractal dimension index of area-weighted mean (FRAC_ AM), and the regional landscape pattern index at the significance level of 0.01. However, the area of Spartina alterniflora was significantly negatively correlated with habitat quality. Overall, the results of this study suggest that the expansion of Spartina alterniflora profoundly affects landscape patterns and functions, warranting control according to local conditions.
XU Ya , ZHANG Huabing . Spatiotemporal trajectory of Spartina alterniflora expansion and its impact on landscape patterns in the Yancheng coastal wetland since the 1980s[J]. Remote Sensing for Natural Resources, 2025 , 37(2) : 246 -255 . DOI: 10.6046/zrzyyg.2023325
表1 互花米草景观格局指数变化Tab.1 Pattern index changes of Spartina alterniflora |
| 年份 | LPI | TE | ED | AI | FRAC_AM | LSI |
|---|---|---|---|---|---|---|
| 1983年 | 1.462 | 20 592.047 | 1.821 | 90.995 | 1.062 | 3.393 |
| 1988年 | 1.011 | 34 792.460 | 3.092 | 85.837 | 1.114 | 5.066 |
| 1992年 | 1.087 | 33 159.686 | 2.934 | 87.507 | 1.104 | 4.810 |
| 1997年 | 6.560 | 40 596.982 | 3.641 | 94.256 | 1.109 | 3.832 |
| 2000年 | 10.874 | 113 108.751 | 10.000 | 92.480 | 1.138 | 6.596 |
| 2002年 | 11.499 | 57 881.943 | 5.117 | 96.978 | 1.103 | 3.359 |
| 2006年 | 25.626 | 86 203.383 | 7.621 | 96.463 | 1.155 | 4.201 |
| 2009年 | 27.190 | 93 283.743 | 8.247 | 96.316 | 1.173 | 4.426 |
| 2011年 | 33.466 | 123 198.264 | 10.892 | 96.048 | 1.157 | 5.137 |
| 2014年 | 30.570 | 91 104.741 | 8.192 | 96.743 | 1.157 | 4.209 |
| 2017年 | 31.674 | 74 166.771 | 6.557 | 97.676 | 1.137 | 3.335 |
| 2021年 | 31.414 | 151 121.920 | 13.325 | 94.422 | 1.202 | 6.480 |
表2 互花米草面积与其景观格局的相关性Tab.2 Correlation between the area and landscape pattern of Spartina alterniflora |
| LPI | TE | ED | AI | FRAC_AM | LSI | |
|---|---|---|---|---|---|---|
| 互花 米草 | 0.982**① | 0.834** | 0.836** | 0.821** | 0.814** | 0.145 |
①“**”表示在0.01水平上显著相关。 |
表3 互花米草扩张对景观结构变化的贡献率Tab.3 Contribution rate of Spartina alterniflora expansion to landscape structure change |
| 时段 | 互花米草扩 张面积/hm2 | 区域景观变 化面积/hm2 | 贡献率/% |
|---|---|---|---|
| 1983—1997年 | 644.604 | 2 461.286 | 26.190 |
| 1997—2006年 | 2 017.240 | 4 807.768 | 41.968 |
| 2006—2021年 | 661.478 | 3 019.392 | 21.908 |
| 1983—2021年 | 3 323.321 | 7 665.844 | 43.352 |
表4 互花米草面积与区域景观格局的相关性Tab.4 Correlation between Spartina alterniflora area and region landscape pattern |
| LPI | TE | ED | AI | FRAC_AM | LSI | SHDI | |
|---|---|---|---|---|---|---|---|
| 互花米草 | -0.914**① | 0.741** | 0.742** | -0.706* | 0.709** | 0.741** | 0.915** |
①“**”表示在0.01水平上显著相关,“*”表示在0.05水平上显著相关。 |
表5 互花米草景观格局与区域景观格局的相关性Tab.5 Correlation between Spartina alterniflora landscape pattern and region landscape pattern |
| 类型 | 区域景观 | ||||||
|---|---|---|---|---|---|---|---|
| LPI | TE | ED | AI | FRAC_AM | LSI | SHDI | |
| LPI | -0.874**① | 0.740** | 0.742** | -0.709** | 0.734** | 0.741** | 0.880** |
| TE | -0.806** | 0.927** | 0.926** | -0.909** | 0.852** | 0.926** | 0.824** |
| ED | -0.808** | 0.927** | 0.926** | -0.909** | 0.853** | 0.926** | 0.827** |
| AI | -0.856** | 0.413 | 0.416 | -0.367 | 0.345 | 0.414 | 0.854** |
| FRAC_AM | -0.811** | 0.871** | 0.871** | -0.855** | 0.880** | 0.871** | 0.815** |
| LSI | -0.165 | 0.641* | 0.638* | -0.659* | 0.625* | 0.640* | 0.185 |
①“**”表示在0.01水平上显著相关,“*”表示在0.05水平上显著相关。 |
| [1] |
张丹华, 胡远满, 刘淼. 基于Maxent生态位模型的互花米草在我国沿海的潜在分布[J]. 应用生态学报, 2019, 30(7):2329-2337.
|
| [2] |
陈正勇, 王国祥, 刘金娥, 等. 苏北潮滩群落交错带互花米草斑块与土著种竞争关系研究[J]. 生态环境学报, 2011, 20(10):1436-1442.
|
| [3] |
袁红伟, 李守中, 郑怀舟, 等. 外来种互花米草对中国海滨湿地生态系统的影响评价及对策[J]. 海洋通报, 2009, 28(6):122-128.
|
| [4] |
|
| [5] |
|
| [6] |
沈鸿坤, 赵博义, 陈铭洋, 等. 1995—2019年广西山口红树林国家级自然保护区互花米草和红树林面积变化[J]. 应用生态学报, 2022, 33(2):397-404.
|
| [7] |
岳英洁, 朱玉玲, 毕永坤, 等. 基于无人机遥感监测的互花米草破碎斑块分布特征研究[J]. 海洋湖沼通报, 2022, 44(4):73-81.
|
| [8] |
|
| [9] |
张华兵, 甄艳, 吴菲儿, 等. 滨海湿地生境质量演变与互花米草扩张的关系——以江苏盐城国家级珍禽自然保护区为例[J]. 资源科学, 2020, 42(5):1004-1014.
|
| [10] |
刘春悦, 张树清, 江红星, 等. 江苏盐城滨海湿地外来种互花米草的时空动态及景观格局[J]. 应用生态学报, 2009, 20(4):901-908.
|
| [11] |
刘永学, 张忍顺, 李满春. 江苏沿海互花米草盐沼动态变化及影响因素研究[J]. 海洋地质动态, 2004, 20(2):18-21,51.
|
| [12] |
郝敬锋, 刘红玉, 李玉凤, 等. 基于转移矩阵模型的江苏海滨湿地资源时空演变特征及驱动机制分析[J]. 自然资源学报, 2010, 25(11):1918-1929.
|
| [13] |
方仁建, 沈永明, 吴德力. 江苏盐城不同沉积岸段湿地景观格局及其时空演变[J]. 生态学杂志, 2014, 33(4):1096-1103.
|
| [14] |
王聪, 刘红玉. 江苏淤泥质潮滩湿地互花米草扩张对湿地景观的影响[J]. 资源科学, 2014, 36(11):2413-2422.
|
| [15] |
张华兵, 刘红玉, 侯明行. 人工管理和自然驱动下盐城海滨湿地互花米草沼泽演变及空间差异[J]. 生态学报, 2013, 33(15):4767-4775.
|
| [16] |
张华兵, 刘红玉, 侯明行, 等. 海滨湿地景观过程模型与模拟研究——以江苏盐城保护区核心区为例[J]. 自然资源学报, 2014, 29(7):1105-1115.
|
| [17] |
王娟, 张华兵, 刘玉卿, 等. 盐城滨海湿地植物群落种间格局与竞争的关系研究[J]. 生态学报, 2020, 40(24):8966-8973.
|
| [18] |
张学勤, 王国祥, 王艳红, 等. 江苏盐城沿海滩涂淤蚀及湿地植被消长变化[J]. 海洋科学, 2006, 30(6):35-39,45.
|
| [19] |
侯明行, 刘红玉, 张华兵, 等. 地形因子对盐城滨海湿地景观分布与演变的影响[J]. 生态学报, 2013, 33(12):3765-3773.
|
| [20] |
宗影, 阳佳伶, 刘红玉, 等. 互花米草入侵对盐城淤泥质潮滩湿地潮沟系统影响研究[J]. 海洋科学进展, 2023, 41(1):109-122.
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
张燕, 孙勇, 鲁长虎, 等. 盐城国家级珍禽自然保护区互花米草入侵后三种生境中越冬鸟类群落格局[J]. 湿地科学, 2017, 15(3):433-441.
|
| [25] |
汪钰明, 高新贻, 谢世君, 等. 互花米草入侵对红树林底栖甲壳动物和鱼类功能群及生态位的影响[J]. 应用生态学报, 2022, 33(11):3016-3026.
|
| [26] |
曾庆淞, 陈钰, 郑毅, 等. 互花米草入侵对河口湿地土壤环境中铁氧化菌群落结构及多样性影响[J]. 环境科学学报, 2022, 42(5):425-434.
|
| [27] |
刘志君, 崔丽娟, 李伟, 等. 互花米草入侵对盐城滨海湿地nirS型反硝化细菌多样性及群落结构的影响[J]. 生态环境学报, 2022, 31(4):704-714.
|
| [28] |
王欢欢, 王银华, 庞金玲, 等. 互花米草入侵背景下增氮对滨海湿地土壤可培养细菌和真菌群落组成及多样性的影响[J]. 南开大学学报(自然科学版), 2022, 55(5):96-105.
|
| [29] |
布乃顺, 胡悦, 杨骁, 等. 互花米草入侵对长江河口湿地土壤理化性质的影响[J]. 长江流域资源与环境, 2017, 26(1):100-109.
|
| [30] |
陈秋宇, 杨仁敏, 朱长明. 基于VIS-NIR光谱的互花米草入侵湿地土壤有机碳预测研究[J]. 土壤学报, 2021, 58(3):694-703.
|
| [31] |
黑杰, 胥佳忆, 王亚非, 等. 互花米草入侵对闽江河口湿地土壤碳、氮、磷及CH4和CO2排放的影响[J]. 环境科学学报, 2022, 42(11):416-426.
|
| [32] |
刘展航, 张树岩, 侯玉平, 等. 互花米草入侵对黄河口湿地土壤碳氮磷及其生态化学计量特征的影响[J]. 生态环境学报, 2022, 31(7):1360-1369.
|
| [33] |
厉彦哲, 孙志高, 毛立, 等. 闽江河口互花米草不同入侵年限湿地土壤磷赋存形态[J]. 应用生态学报, 2022, 33(4):1003-1011.
|
| [34] |
张治梅, 樊彦国, 矫志军, 等. 土壤盐渍化对滨海湿地生态环境质量的影响——以黄河三角洲为例[J]. 自然资源遥感, 2023, 35(4):226-235.doi:10.6046/zrzyyg.2022284.
|
| [35] |
钦佩. 互花米草与人体健康关系研究进展[J]. 中国野生植物资源, 2019, 38(5):70-73.
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}