
Effects of environmental factors on anthocyanin accumulation in Lycium ruthenicum from various provenances
Received date: 2025-01-22
Revised date: 2025-05-30
Online published: 2026-03-12
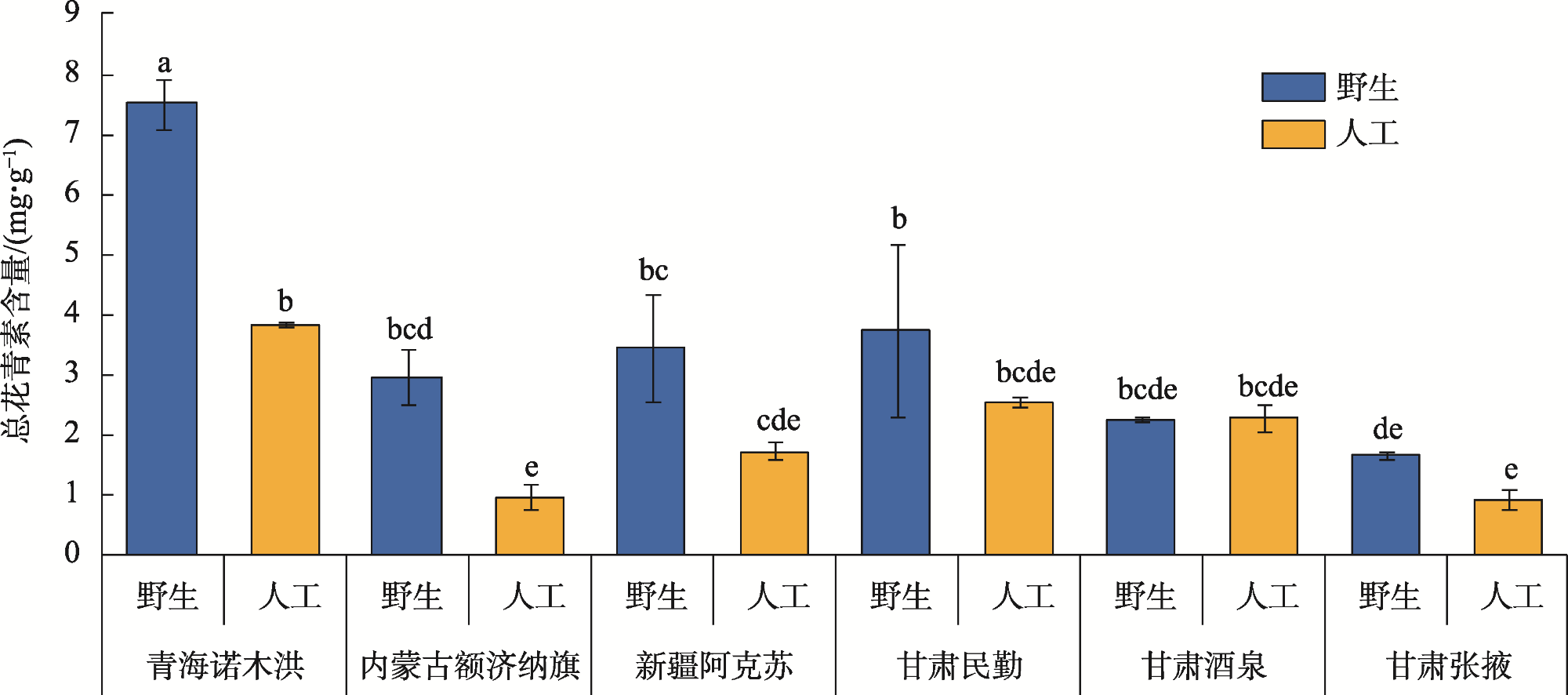
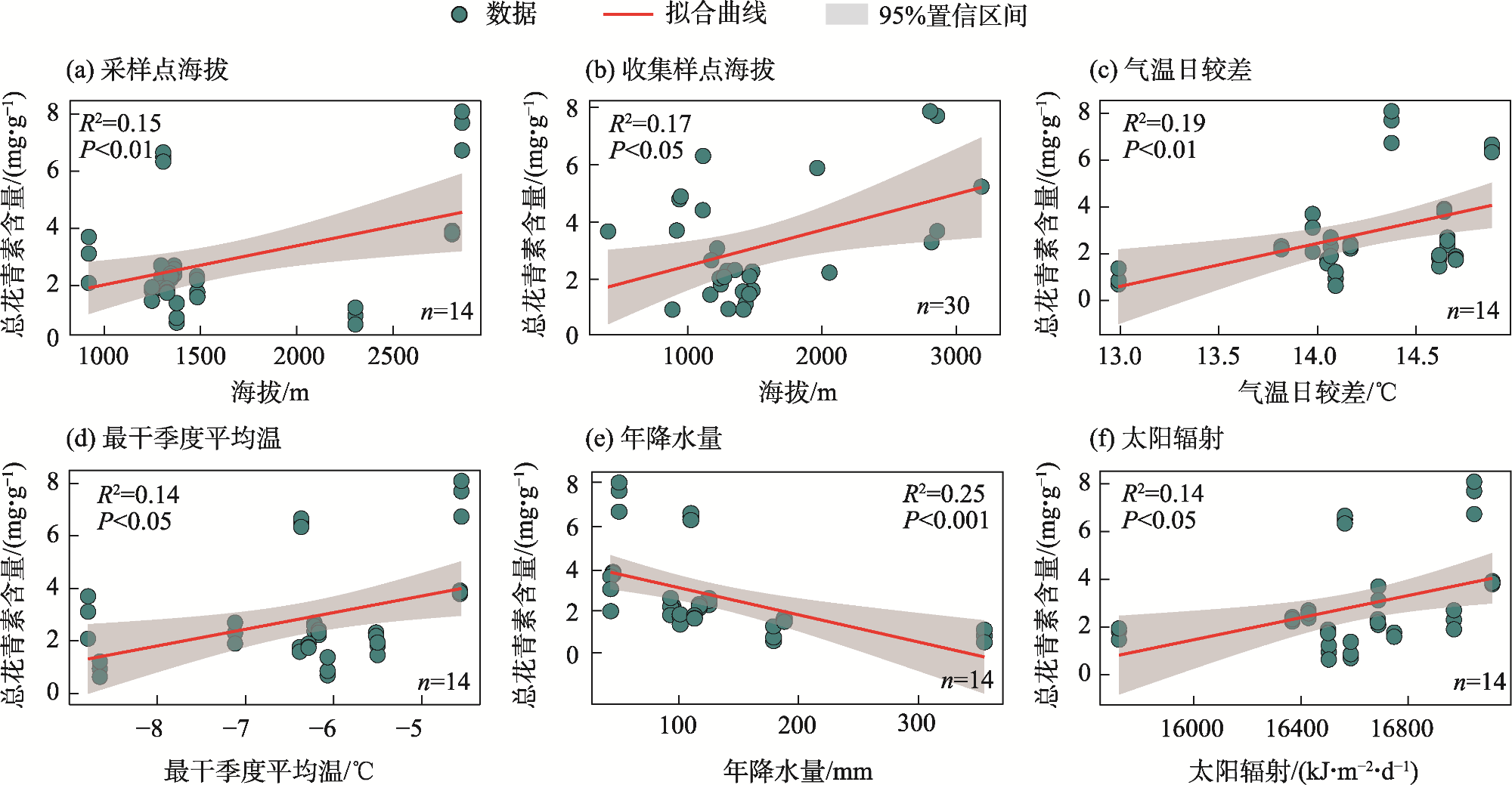
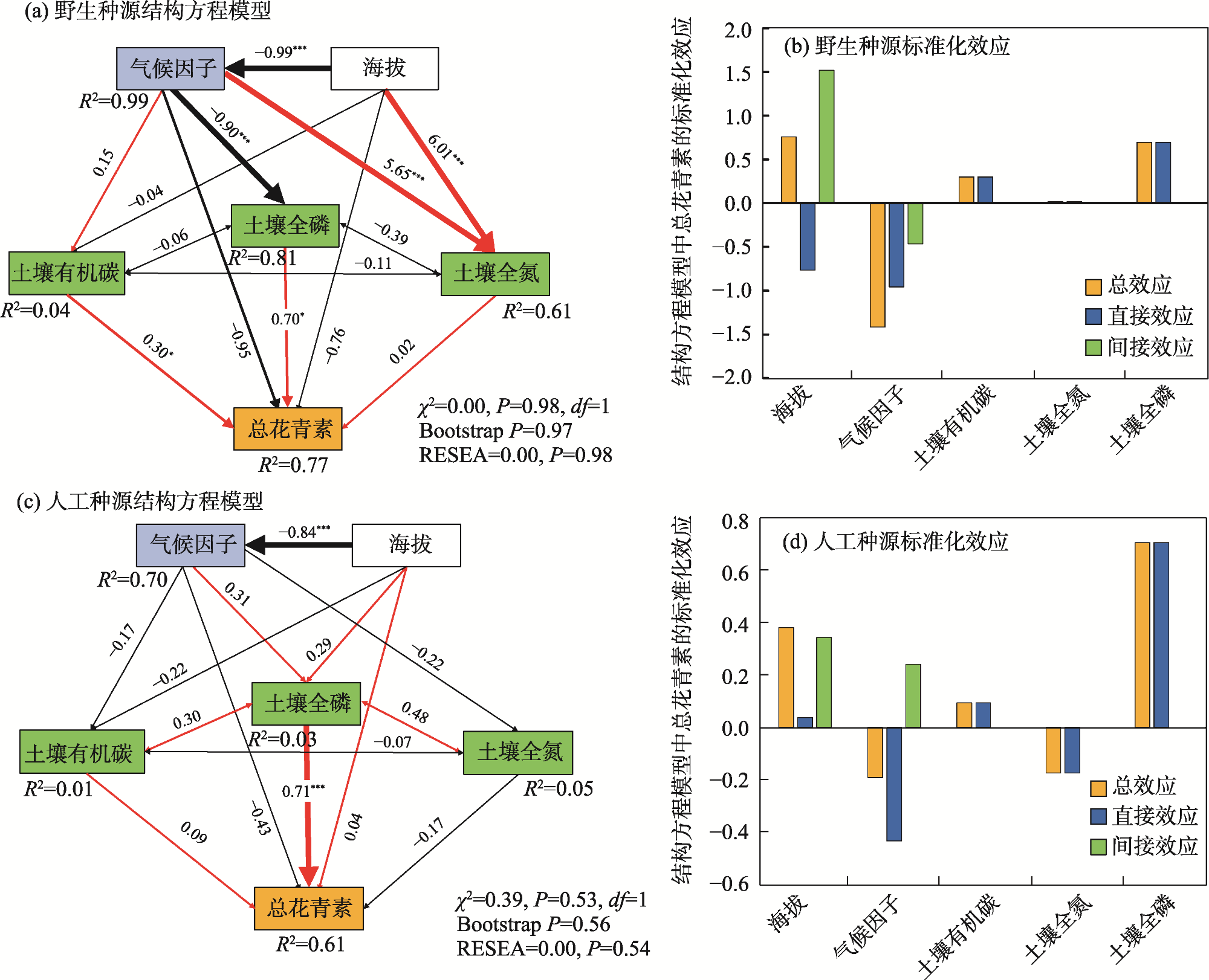
Anthocyanins are potent free radical scavengers that help plants adapt to extreme environmental stresses and have important nutritional and medicinal value for humans. Studying the effect of habitat conditions on the variation in germplasm in Lycium ruthenicum can provides an important scientific basis for artificial breeding and germplasm innovation. L. ruthenicum collected from various provenances was used as the research object. Analytical methods, including structural equation model (SEM), principal component analysis, and linear regression analysis, were used to explore the effects of space, climate, and soil factors on anthocyanin accumulation in L. ruthenicum fruits. The results showed that: (1) The accumulation of anthocyanin in L. ruthenicum fruits increased significantly as altitude increased (P<0.01); altitude mainly influenced climate (β=−0.99, P<0.001) and soil total phosphorus (β=−0.90, P<0.001). (2) The accumulation of anthocyanin in wild L. ruthenicum fruits was mainly influenced by climatic factors, and the artificial cultivation of L. ruthenicum fruits was mainly influenced by soil total phosphorus. (3) The important climatic factors influencing anthocyanin accumulation in L. ruthenicum were diurnal temperature range (DTR), mean temperature of driest quarter (MTDQ), mean annual precipitation (MAP), and solar radiation. Therefore, environmental factors influencing the anthocyanin accumulation in L. ruthenicum depend on the provenance. Thus, when regulating and improving anthocyanin accumulation, it is necessary to adjust measures to local conditions and develop improvement measures to suit the corresponding environmental driving factors.
LI Jinhui , HU Jing , JIN Hongxi , WANG Qi , YAO Ze . Effects of environmental factors on anthocyanin accumulation in Lycium ruthenicum from various provenances[J]. Arid Zone Research, 2025 , 42(8) : 1525 -1535 . DOI: 10.13866/j.azr.2025.08.15
表1 14个采样点的地理、气候信息Tab. 1 Geographic, climatic information of the fourteen sampling sites |
| 采样点 | 经度(E) | 纬度(N) | 海拔/m | 气温日较差/℃ | 最干季度平均温/℃ | 年降水量/mm | 太阳辐射/(kJ·m-2·d-1) |
|---|---|---|---|---|---|---|---|
| 青海诺木洪-野生 | 96°11′ | 36°30′ | 2858 | 14.37 | -4.55 | 50 | 17047.00 |
| 青海诺木洪-人工 | 96°11′ | 36°23′ | 2806 | 14.64 | -4.57 | 45 | 17114.75 |
| 内蒙古额济纳旗-野生 | 101°10′ | 41°59′ | 919 | 13.97 | -8.78 | 43 | 16688.83 |
| 内蒙古额济纳旗-人工 | 105°32′ | 38°47′ | 1376 | 12.99 | -6.07 | 179 | 16585.92 |
| 新疆阿克苏-野生 | 79°10′ | 41°08′ | 1396 | 12.13 | -4.55 | 125 | 15676.67 |
| 新疆阿克苏-人工 | 80°56′ | 41°26′ | 1247 | 14.62 | -5.50 | 101 | 15720.17 |
| 甘肃民勤-野生 | 103°31′ | 38°46′ | 1345 | 14.17 | -6.17 | 117 | 16367.75 |
| 甘肃民勤-野生 | 102°57′ | 38°50′ | 1326 | 14.70 | -6.28 | 113 | 16501.42 |
| 甘肃民勤-野生 | 103°36′ | 39°03′ | 1306 | 14.88 | -6.37 | 110 | 16563.92 |
| 甘肃民勤-人工 | 102°58′ | 38°35′ | 1363 | 14.66 | -6.22 | 125 | 16428.17 |
| 甘肃酒泉-野生 | 98°27′ | 39°56′ | 1481 | 13.82 | -5.52 | 95 | 16687.00 |
| 甘肃酒泉-人工 | 98°53′ | 39°55′ | 1295 | 14.07 | -7.12 | 93 | 16971.42 |
| 甘肃张掖-野生 | 100°25′ | 39°01′ | 1483 | 14.05 | -6.38 | 188 | 16748.00 |
| 甘肃张掖-人工 | 99°35′ | 38°49′ | 2304 | 14.09 | -8.65 | 355 | 16503.75 |
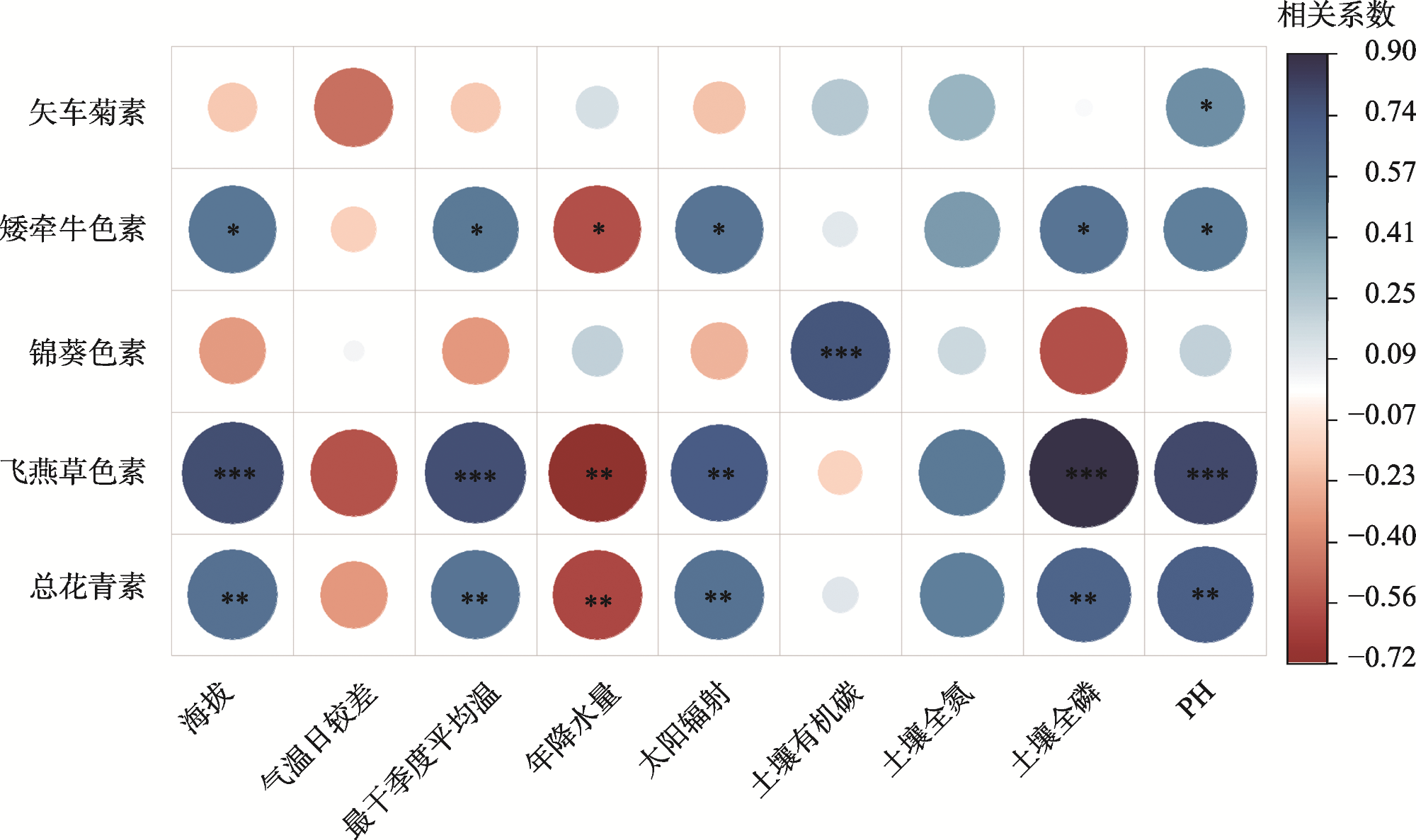
图3 不同种源野生和人工种植黑果枸杞生境海拔-气候-土壤影响总花青素积累的结构方程模型注:系数是对每条因果路径的标准化预测;红色实线箭头表示正效应;黑色实线箭头表示负效应;箭头的粗细与标准化路径系数的大小成正比,表明关系的强弱;*和***分别表示在P<0.05和P<0.001水平显著相关;χ2表示卡方值;df表示自由度;RMSEA表示均方根近似误差;R2表示决定系数。 Fig. 3 SEM of altitude-climate-soil effects on total anthocyanin accumulation in wild and cultivated Lycium ruthenicum habitat from various provenances |
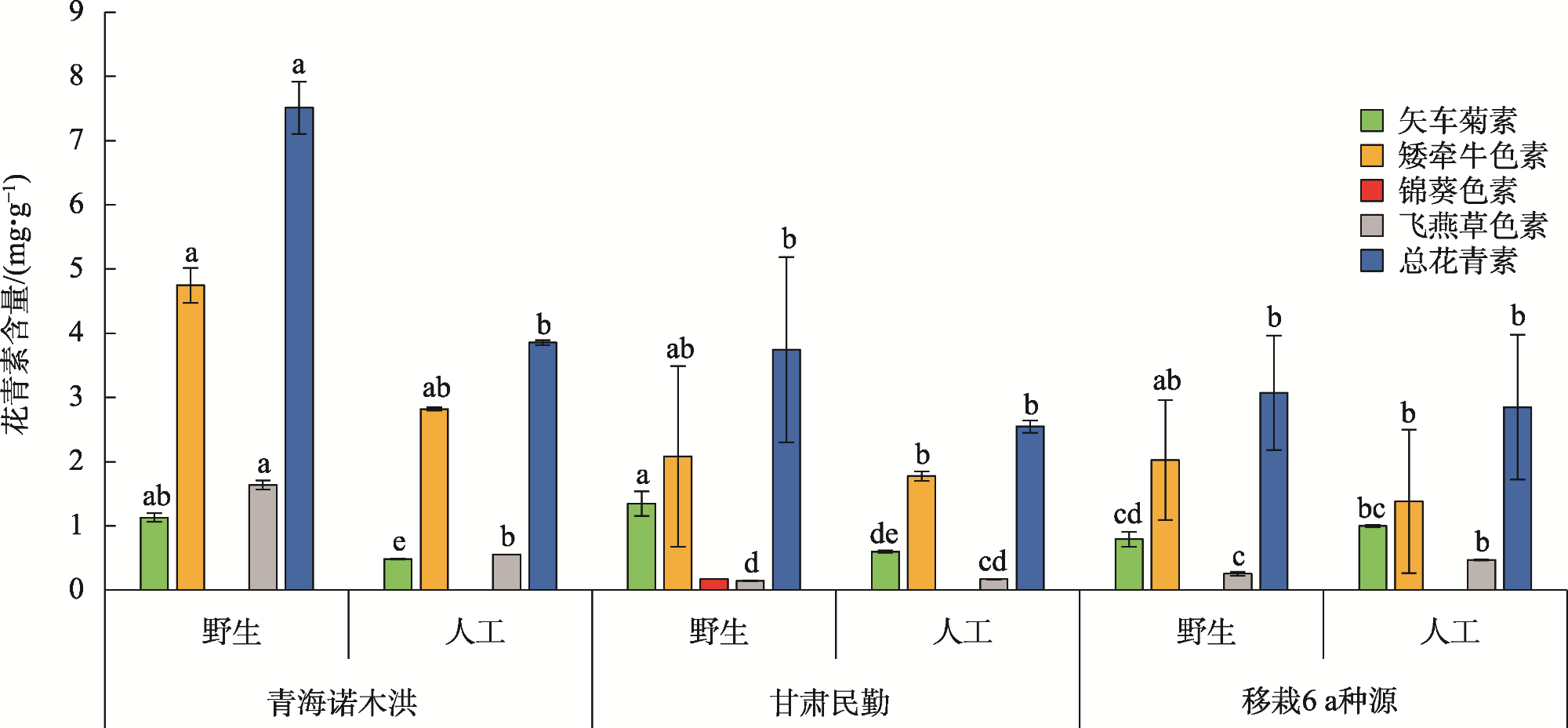
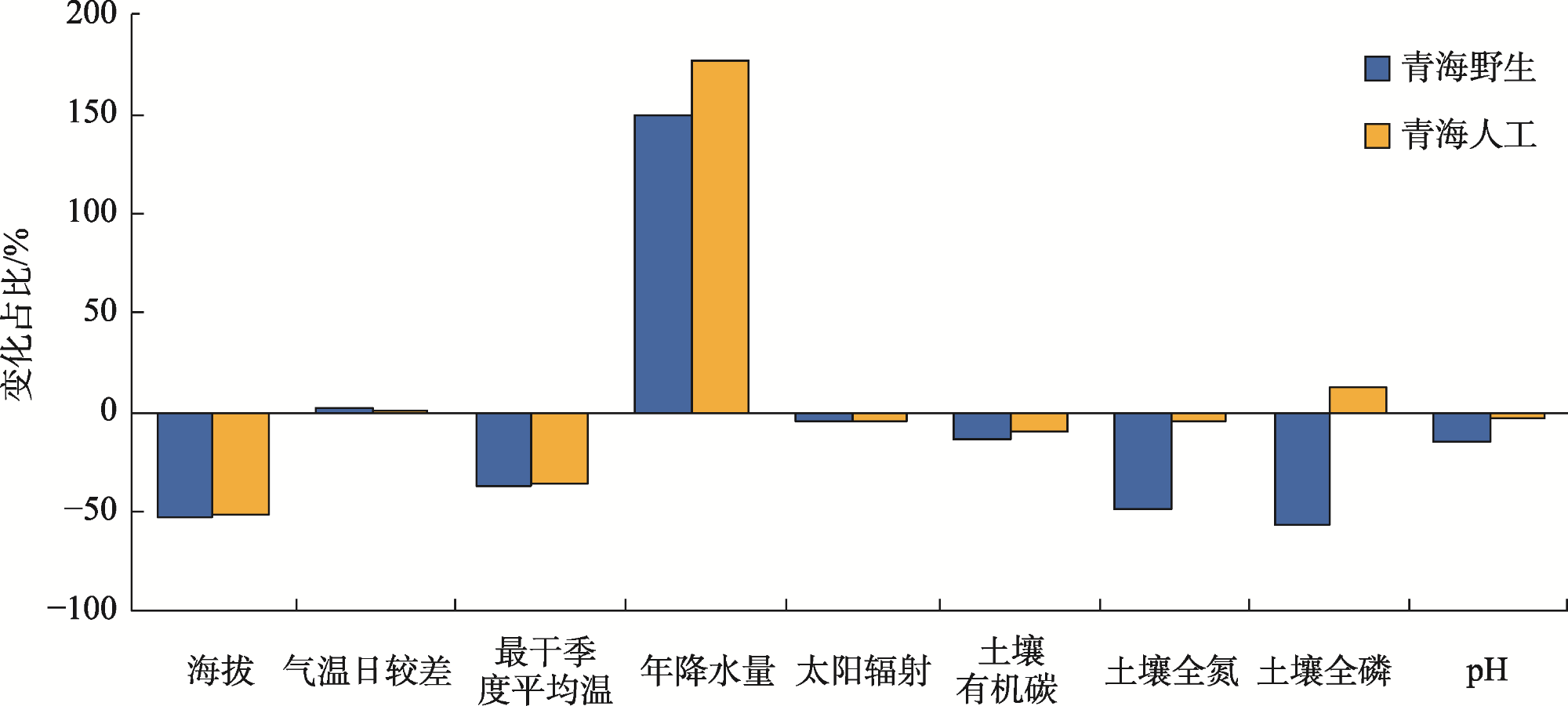
图5 青海诺木洪种源黑果枸杞移栽民勤后环境因子变化占比Fig. 5 Proportion of environmental factors changes after transplanting Lycium ruthenicum from Qinghai nuomuhong to Minqin |
本研究野外调查、样地设置和采样过程中雷成和给予了很大的帮助,在此表示衷心的感谢!
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
魏永赞, 胡福初, 郑雪文, 等. 光照对荔枝果实着色和花色素苷生物合成影响的分子机制研究[J]. 园艺学报, 2017, 44(7): 1363-1370.
[
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
董鹏, 任悦, 高广磊, 等. 呼伦贝尔沙地樟子松枯落物和土壤碳, 氮, 磷化学计量特征[J]. 干旱区研究, 2024, 41(8): 1354-1363.
[
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
蒋宝, 蒲飞, 孙占育, 等. 海拔对酿酒葡萄果实和相应葡萄酒中多酚物质影响的研究概述[J]. 食品与发酵工业, 2016, 42(8): 262-267.
[
|
| [19] |
矫晓丽, 迟晓峰, 董琦, 等. 柴达木野生黑果枸杞营养成分分析[J]. 氨基酸和生物资源, 2011, 33(3): 60-62.
[
|
| [20] |
彭飞, 黄翠华, 尤全刚, 等. 种植黑果枸杞(Lycium ruthenicum)对盐渍土盐分分布的影响[J]. 中国沙漠, 2013, 33(5): 1406-1412.
[
|
| [21] |
|
| [22] |
|
| [23] |
刘克彪, 郭春秀, 张元恺, 等. 不同种源黑果枸杞物候期和生长差异及其与地理-气候因子的相关性分析[J]. 植物资源与环境学报, 2019, 28(4): 41-48.
[
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
李金霞, 朱亚男, 孙小妹, 等. 氮磷添加对黑果枸杞(Lycium ruthenicum)营养器官非结构性碳水化合物特征的影响[J]. 中国沙漠, 2021, 41(2): 200-211.
[
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
热依拉穆·麦麦提吐尔逊, 哈里布努尔, 艾沙江·阿不都沙拉木. 异质生境下黑果枸杞异形果实的种子休眠及萌发特性[J]. 干旱区研究, 2023, 40(7): 1152-1163.
[
|
| [37] |
|
| [38] |
|
| [39] |
何彩, 戴建昊, 刘伟, 等. 荒漠区黑果枸杞造林及人工栽培种源筛选[J]. 经济林研究, 2021, 39(2): 90-96.
[
|
| [40] |
|
| [41] |
|
| [42] |
李发奎, 李金霞, 孙小妹, 等. 黑果枸杞茎叶生长及其生态化学计量特征对灌水施肥的响应[J]. 干旱区研究, 2020, 37(2): 452-461.
[
|
| [43] |
|
| [44] |
|
| [45] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}