
Analysis of the expression patterns of the heat-resistant gene of Syntrichia caninervis under different abiotic stresses based on RT-qPCR technology
Received date: 2025-01-10
Revised date: 2025-03-26
Online published: 2026-03-12
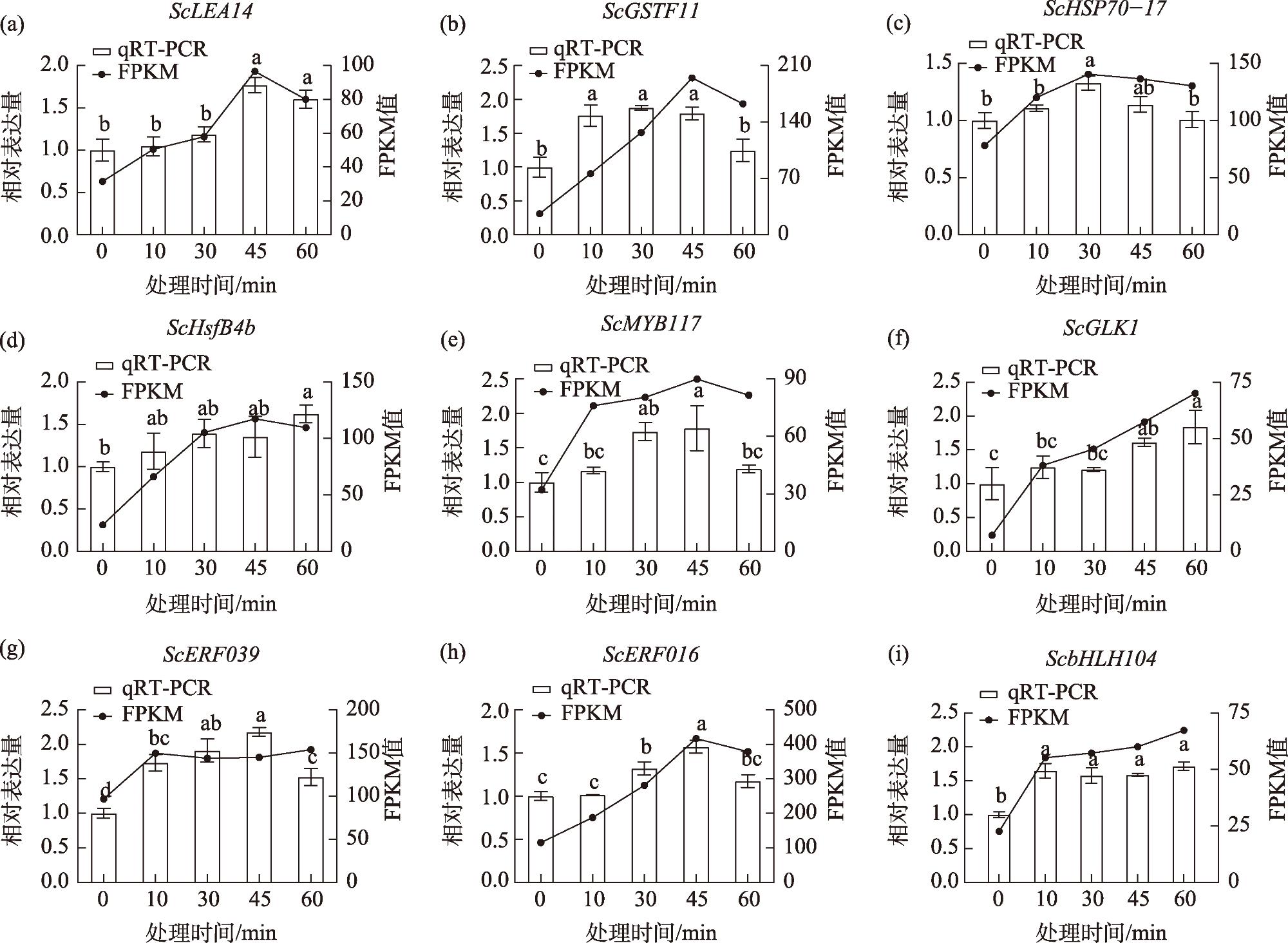
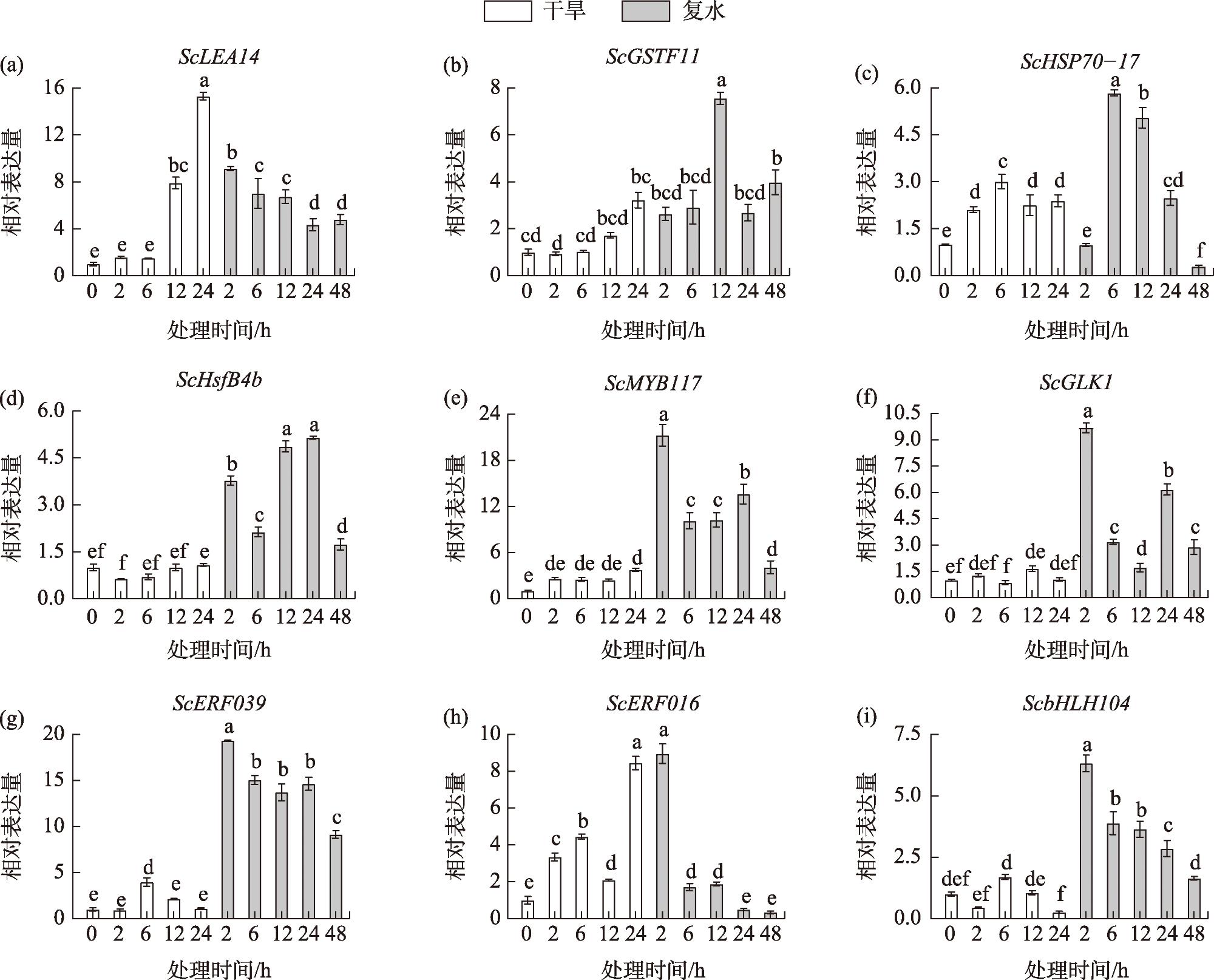
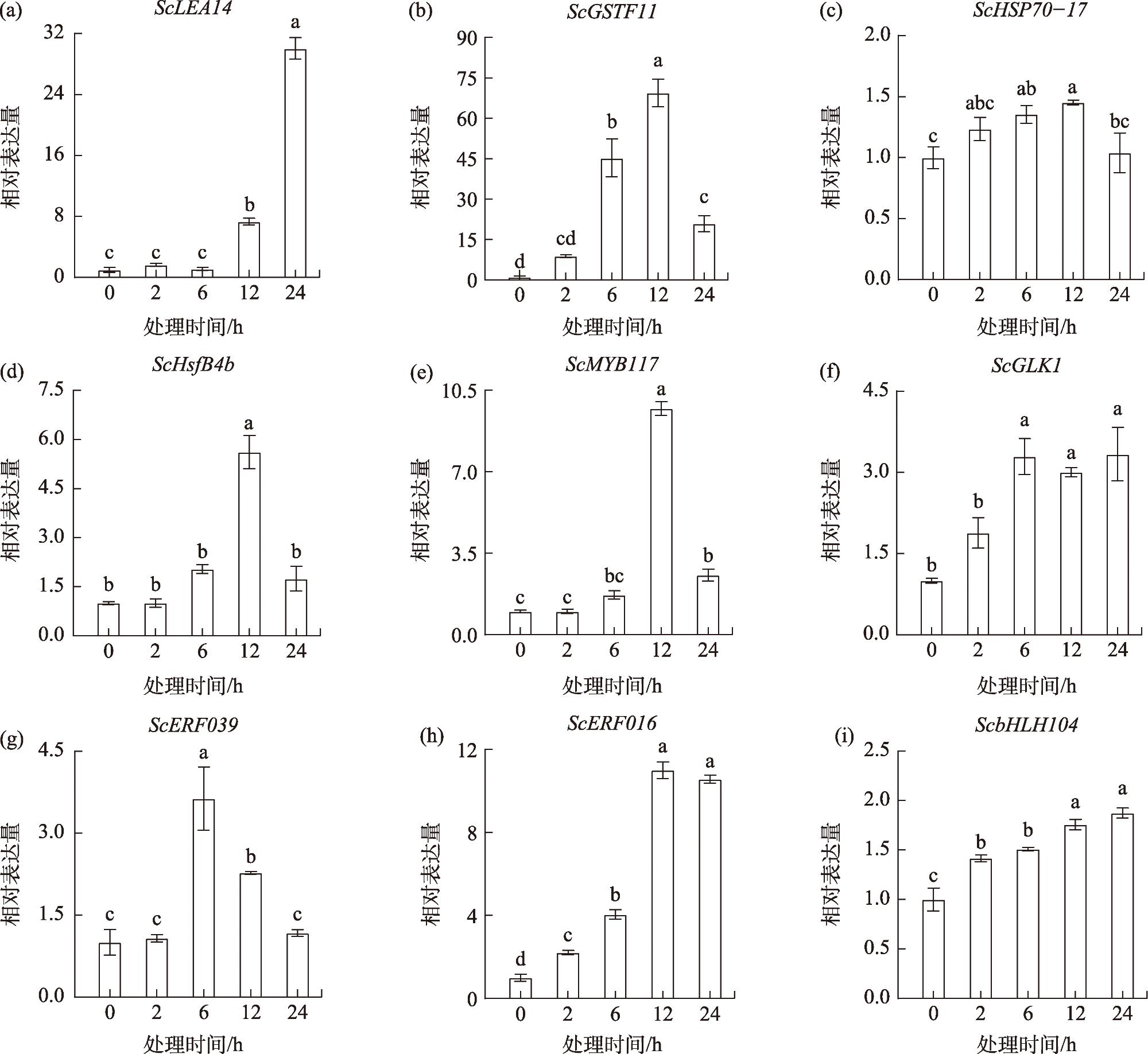
Based on transcriptomic data of the desiccation-tolerant moss Syntrichia caninervis under prior 55 ℃ heat stress, this study employed real-time fluorescent quantitative PCR (RT-qPCR) to investigate the expression patterns of nine heat-responsive differentially expressed genes (ScLEA14, ScGSTF11, ScHSP70-17, ScHsfB4b, ScMYB117, ScGLK1, ScERF039, ScERF016, and ScbHLH104) under high temperature, drought-rehydration, and NaCl stress conditions. The aim was to validate the reliability of RNA-Seq data and provide theoretical support for subsequent functional studies on stress-resistant genes in S. caninervis. Results demonstrated that: (1) The expression profiles of all nine genes under high-temperature stress exhibited substantial concordance with RNA-Seq data, confirming the stability of transcriptomic sequencing. (2) Under extreme heat and drought-rehydration stresses, all genes were differentially induced, with three genes attaining peak expression levels following 24-hour drought treatment, while eight genes displayed more prominent transcriptional activation during the rehydration phase. (3) NaCl stress triggered significant upregulation of all nine thermotolerance-associated genes, with six genes demonstrating statistically robust induction. Thus, the results demonstrate that the three genes ScLEA14, ScMYB117, and ScERF016 are strongly induced under extreme high temperature, drought-rehydration, and NaCl-induced high salinity stress, highlighting their potential as key candidate genes for further investigation into stress resistance mechanisms.
HUO Wenting , GU Tianqi , GAO Mengyu , SONG Yanfang , LI Hongbin , ZHUO Lu . Analysis of the expression patterns of the heat-resistant gene of Syntrichia caninervis under different abiotic stresses based on RT-qPCR technology[J]. Arid Zone Research, 2025 , 42(5) : 885 -894 . DOI: 10.13866/j.azr.2025.05.11
表1 齿肋赤藓9个耐热相关基因实时荧光定量PCR引物序列信息Tab. 1 Nine heat tolerance related genes primer sequences information for RT-qPCR analysis of Syntrichia caninervis |
| 基因 | 引物序列(5′→3′) | 产物长度/bp | |
|---|---|---|---|
| 正向引物 | 反向引物 | ||
| ScLEA14 | ATGATCACGATCTGCCCAT | TTATGCGGCTCCCAACTGA | 148 |
| ScGSTF11 | GAGTCTCACCAAGTTAAACAGCC | ATCTTCTTTTCCACGTACTGCAA | 89 |
| ScHSP70-17 | GAAGGACTGAGCATGTTTCCTCT | AGGCCATCTCTGTACAATCAGC | 150 |
| ScHsfB4b | CAGTACCAGTTCAAAGGATCACCC | TTCGTTTGGACAATGCAGTCACT | 84 |
| ScMYB117 | CGGTGCTGCTCTGAAACCAA | CTTGCCTGAAGCACAAACCAC | 82 |
| ScGLK1 | CCTCCTCCGACCTGTTAGATGT | CGCAACTAATCCCCGATGCC | 129 |
| ScERF039 | GTTGCATCTGCCGTCATCGAAC | CCCGCTACTACTGTCCTTCGTC | 118 |
| ScERF016 | AGAAGAATGGGAGTCAGAGAGAAGC | CCTTCACGCTCTTCTTCGCTAC | 81 |
| ScbHLH104 | TCACAACTGCGGTATCTGGT | AATCGTAGCTTTGTCGGTCT | 125 |
| α-TUB | CGGTCATTACACCGTGGGAA | CCTCTCCAGCAACAGCGAA | 243 |
表2 基于RNA-Seq数据筛选的齿肋赤藓耐热相关响应基因Tab. 2 Heat tolerance related genes based on RNA-Seq technique of Syntrichia caninervis |
| 基因分类 | 基因名称 | 基因差异表达倍数变化(Log2) | |||
|---|---|---|---|---|---|
| 10 min | 30 min | 45 min | 60 min | ||
| 功能基因 | ScLEA14 | +0.73 | +0.98 | +1.71 | +1.37 |
| ScGSTF11 | +1.59 | +2.39 | +2.99 | +2.66 | |
| ScHSP70-17 | +0.67 | +0.95 | +0.90 | +0.77 | |
| 转录因子 | ScHsfB4b | +1.54 | +2.27 | +2.42 | +2.25 |
| ScMYB117 | +1.28 | +1.43 | +1.58 | +1.37 | |
| ScGLK1 | +2.45 | +2.76 | +3.10 | +3.31 | |
| ScERF039 | +0.67 | +0.68 | +0.68 | +0.70 | |
| ScERF016 | +0.76 | +1.39 | +1.95 | +1.75 | |
| ScbHLH104 | +1.33 | +1.45 | +1.51 | +1.60 | |
注:+为表达上调。 |
图1 齿肋赤藓9个耐热相关基因在高温处理下实时荧光定量PCR分析和RNA-Seq基因表达谱注:FPKM是指每百万个比对上的reads中比对到外显子的每1000个碱基上的片段个数;相同小写字母为差异不显著,不同小写字母为差异显著,小写字母表示显著水平α=0.05。下同。 Fig. 1 Quantitative real-time PCR analysis and RNA-Seq gene expression profiles of nine heat-tolerance-associated genes under high-temperature treatment of Syntrichia caninervis |
| [1] |
陈颖, 邵伟玲, 曹萌, 等. 新疆夏季高温日数的变化特征及其影响因子[J]. 干旱区研究, 2020, 37(1): 58-66.
[
|
| [2] |
刘璐, 刘普幸, 张旺雄, 等. 1961—2017年新疆极端暖事件变化特征及其未来情景预估[J]. 干旱区研究, 2021, 38(6): 1590-1600.
[
|
| [3] |
林晓华, 卓一林, 柳丽芳, 等. NaCl胁迫对荒漠苔藓齿肋赤藓膜结构稳定性的影响[J]. 生态学报, 2024, 44(8): 3483-3491.
[
|
| [4] |
|
| [5] |
聂华丽, 吴楠, 梁少民, 等. 不同沙埋深度对刺叶墙藓植株碎片生长的影响[J]. 干旱区研究, 2006, 23(1): 66-70.
[
|
| [6] |
马瑞. 荒漠齿肋赤藓抗旱基因ScALDH21、Sc288对干旱胁迫的响应[D]. 长沙: 中南林业科技大学, 2011.
[
|
| [7] |
杨红兰, 张道远, 刘燕, 等. 齿肋赤藓乙醛脱氢酶基因ALDH21的克隆与表达分析[J]. 基因组学与应用生物学, 2010, 29(1): 24-30.
[
|
| [8] |
|
| [9] |
韩志立, 尹本丰, 杨孜悦, 等. 积雪变化对温带荒漠齿肋赤藓结皮土壤磷组分的影响[J]. 生态学报, 2024, 44(16): 7119-7129.
[
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
薛山. 新疆特色沙漠植物齿肋赤藓热胁迫响应关键基因的筛选与分析[D]. 石河子: 石河子大学, 2024.
[
|
| [22] |
梁玉青, 李小双, 高贝, 等. 基于RNA-Seq数据筛选的银叶真藓耐干相关基因表达模式研究[J]. 植物生理学报, 2017, 53(3): 388-396.
[
|
| [23] |
|
| [24] |
宋士伟, 焦德志, 陈旭, 等. 野大麦对干旱胁迫的生理响应与转录组分析[J]. 干旱区研究, 2019, 36(4): 909-915.
[
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
黄畋柳, 张锐, 贺迎骁, 等. 辣椒NAC家族成员鉴定及其编码基因在NaCl胁迫下的表达分析[J]. 植物资源与环境学报, 2023, 32(4): 12-24.
[
|
| [35] |
|
| [36] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}